Двустворчатые
| Двустворчатые | |||||||||||||||||
 «Acephala», иллюстрация из работы Э. Геккеля «Красота форм в природе» (1904) | |||||||||||||||||
| Научная классификация | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
промежуточные ранги
| |||||||||||||||||
| Международное научное название | |||||||||||||||||
Bivalvia Linnaeus, 1758 | |||||||||||||||||
| Синонимы | |||||||||||||||||
| |||||||||||||||||
| Подклассы | |||||||||||||||||
| |||||||||||||||||
| |||||||||||||||||
Двуство́рчатые, или пластинчатожа́берные (лат. Bivalvia) — класс морских и пресноводных малоподвижных моллюсков, тело которых уплощено с боков и заключено в раковину из двух створок[1]. К ним относят таких известных моллюсков, как устрицы, мидии, морские гребешки. В отличие от всех других моллюсков Bivalvia лишены головы и радулы. Большинство представителей класса имеют сильно развитые пластинчатые жабры (ктенидии), выполняющие не только дыхательную функцию, но и роль фильтров для отцеживания пищевых частиц из воды, поэтому по типу питания двустворчатые — преимущественно фильтраторы. Большинство двустворчатых закапываются в донный ил, спасаясь таким образом от хищников, некоторые лежат на морском дне или облепляют скалы и другие поверхности. Немногие виды, например морские гребешки, способны к непродолжительному активному плаванию. В целом же особенности внешнего и внутреннего строения двустворчатых отражают их экологическую специализацию к малоподвижному или неподвижному образу жизни[2].
Раковины двустворчатых моллюсков состоят из карбоната кальция и представляют собой две обычно равные по величине створки, скреплённые друг с другом с одного края, возле макушки, упругой белковой связкой — лигаментом. Кроме того, их соединение обеспечивают чередующиеся выступы (зубы) и выемки, расположенные на каждой створке раковины и составляющие замок. Такое устройство раковины позволяет открывать створки для питания или передвижения и плотно закрывать их в случае опасности. Как правило, раковина двусторонне-симметричная. Её размер варьирует от нескольких миллиметров до метра и более, однако у большинства представителей он не превышает 10 см.
Начиная с доисторического времени, двустворчатые моллюски составляли важную часть рациона жителей побережья. Ещё римляне разводили устриц в прудах, а ныне большинство употребляемых в пищу двустворчатых моллюсков выращивают в марикультуре. Изучение жизненных циклов съедобных двустворчатых сделало возможным разведение их в питомниках, а также разработку новых технологий их разведения. Помимо пищевого применения, Bivalvia являются производителями натурального жемчуга. Жемчужины используют в ювелирном деле, а перламутр из раковин двустворчатых — в изготовлении пуговиц или дешёвых украшений. Двустворчатые также могут служить биоиндикаторами загрязнения окружающей среды. Наконец, они представляют собой ценный корм для рыб и других животных[2].
Находки древнейших ископаемых двустворчатых моллюсков датируются началом кембрийского периода, их возраст составляет более 500 млн лет. Общее число ныне живущих видов составляет приблизительно 9200[3] (по другим данным, более 20 тысяч[2]). Их включают в 1260 родов и 106 семейств[3][4].
В России встречается около 1000 видов двустворчатых[5]. Морские пластинчатожаберные (включая виды, обитающие в солоноватых и эстуарных водах) представлены примерно 8000 видами, которые включают в 4 подкласса, 99 семейств и 1100 родов. Крупнейшие семейства пластинчатожаберных — венериды (Veneridae; более 680 видов), а также теллиниды (Tellinidae) и люциниды (Lucinidae), каждое из которых содержит более 500 видов. Пресноводные Bivalvia составляют 7 семейств, крупнейшее из которых — униониды (Unionidae) — включает около 700 видов. Хотя общим числом видов двустворчатые в несколько раз уступают брюхоногим, по численности и биомассе на единицу площади морского дна они значительно превосходят другие группы моллюсков[2].
Содержание
1 Этимология
2 Ареал и местообитание
3 Строение и физиология
3.1 Внешнее строение
3.1.1 Размеры и форма тела
3.1.2 Раковина
3.1.3 Мантия и мантийная полость
3.1.4 Нога
3.2 Внутреннее строение
3.2.1 Мускулатура
3.2.2 Пищеварительная система
3.2.3 Нервная система
3.2.4 Органы чувств
3.2.5 Дыхательная система
3.2.6 Кровеносная система
3.2.7 Выделительная система
3.2.8 Осморегуляция
3.2.9 Половая система
3.3 Отличия от плеченогих
4 Жизненный цикл
5 Питание
6 Образ жизни
7 Экология
7.1 Роль в экосистемах
7.2 Взаимоотношения с другими организмами
7.2.1 Естественные враги
7.2.2 Симбионты
8 Эволюция
9 Классификация
9.1 Практическая классификация Р. К. Мура
9.2 Классификация 1935 года
9.3 Классификация на основе строения замка
9.4 Классификация на основе строения жабр
9.5 Классификация 2010 года
10 Разнообразие двустворчатых
11 Охрана двустворчатых
12 Взаимодействие с человеком
12.1 Использование человеком
12.2 Вредители
12.3 Двустворчатые и здоровье человека
13 Двустворчатые моллюски в культуре
14 Примечания
15 Литература
15.1 На русском языке
15.2 На английском языке
Этимология |
Впервые название Bivalvia употребил Карл Линней в 10-м издании своего труда «Система природы» в 1758 году для обозначения моллюсков, раковина которых состоит из двух створок[6]. Кроме того, этот класс известен под названиями Pelecypoda (что означает «топороногие»), Lamellibranchia (пластинчатожаберные) и Acephala (так как двустворчатые, в отличие от всех других моллюсков, утратили голову)[7].
Слово Bivalvia образовано от лат. bis — «два» и лат. valvae — «створка»[7]. Однако не все животные, имеющие раковину из двух створок, относятся к двустворчатым моллюскам. Так, раковину из двух створок имеют брюхоногие моллюски семейства Juliidae[8], представители типа плеченогие (Brachiopoda)[9], а также ракообразные класса ракушковые (Ostracoda)[10] отряда Cyclestherida[11].
В российской науке конца XVIII, начала XIX веков двустворчатые (живые и ископаемые) назывались — черепнокожие[12].
Ареал и местообитание |

Антарктический гребешок Adamussium colbecki под 5-метровым слоем морского льда в море Росса, Антарктида
Двустворчатые моллюски — довольно успешный класс беспозвоночных, исключительно водных и встречающихся в пресных и солёных водах по всему миру. Большинство являются бентосными организмами и живут, зарываясь в донные отложения или прикрепляясь к подводным предметам. Многие двустворчатые освоили литораль и сублиторальные зоны океанов. Хотя песчаный пляж может на первый взгляд показаться лишённым жизни, в песке всегда можно найти много двустворчатых моллюсков и других беспозвоночных. На одном крупном пляже в Южном Уэльсе количество двустворчатых моллюсков сердцевидка съедобная (Cerastoderma edule) оценивают как 3,55 млн особей на гектар[13].
Двустворчатые населяют как тропики, так и умеренные и арктические воды. Некоторые виды могут выживать и даже процветать в экстремальных условиях. Двустворчатые многочисленны в Арктике, где известно около 140 их видов[14]. Антарктический гребешок Adamussium colbecki обитает под слоем морского льда при температурах ниже нуля, когда скорость обмена веществ очень низка[15]. Моллюски Bathymodiolus thermophilus и Calyptogena magnifica образуют скопления вокруг гидротермальных глубоководных источников в Тихом океане на глубине около 3 км[16]. Они вступают в симбиоз с хемосинтезирующими бактериями, окисляющими сероводород, и питаются синтезируемыми ими питательными веществами[17]. Морскую устрицу Enigmonia aenigmatica можно назвать земноводным моллюском. Она обитает в тропической зоне Индийского и Тихого океанов на ветвях и обратной стороне листьев мангровых деревьев, а также на волнорезах[16].
Некоторые пресноводные формы имеют крайне ограниченный ареал. Например, Villosa arkansasensis обитает исключительно в ручьях гор Уошито в Арканзасе и Оклахоме и, наряду с несколькими другими пресноводными двустворчатыми юго-востока США, находится под угрозой исчезновения[18]. Часть пресноводных двустворчатых, напротив, распространилась чрезвычайно широко, например Limnoperna fortunei. Этот вид увеличил свой ареал от Юго-Восточной Азии до Аргентины, где стал инвазивным видом[19]. Другой пресноводный двустворчатый моллюск, речная дрейссена (Dreissena polymorpha), изначально обитавшая на юго-востоке России, была случайно интродуцирована во внутренние воды Северной Америки и Европы, где этот вид нанёс вред находящимся в воде сооружениям и разрушил местные экосистемы[20].
Строение и физиология |
Внешнее строение |
Тело двустворчатых моллюсков, как правило, состоит из туловища, заключающего внутренние органы, и мускулистой ноги, в то время как голова редуцирована. Тело прикрыто раковиной из двух створок, выделяемых складками мантии. Нога, как правило, клиновидная, но у неподвижных форм редуцируется[21].
Размеры и форма тела |

Гигантская тридакна (Tridacna gigas)

Морской черенок (Ensis ensis)
Форма и размер раковин двустворчатых сильно варьируют[21]. К классу двустворчатых относят самых маленьких по размеру представителей типа моллюсков. Взрослые особи наиболее мелкого вида, Condylonucula maya, достигают в длину лишь 0,5 мм[16]. Гигант среди двустворчатых моллюсков — гигантская тридакна (Tridacna gigas) — может достигать 1,4 м в длину и весить до 200 кг[21]. Наибольшая длина тела у корабельного червя Kuphus polythalamia — 1,532 м[22]. Крупнейшим ископаемым моллюском считается Platyceramus, чьи останки достигают до 3 м в длину[23].
Форма тела у двустворчатых тоже весьма различна. Например, сердцевидки имеют почти шаровидное тело и могут прыгать, сгибая и разгибая ногу. В то же время морские черенки (Ensis) в связи со специализацией к роющему образу жизни имеют продолговатую раковину и сильную ногу, предназначенную для закапывания в грунт. Корабельные черви семейства Teredinidae — сильно удлинённое червеобразное тело с редуцированной раковиной, располагающейся на его переднем конце и видоизменённой в сверлильный орган, благодаря которому моллюск «прогрызает» разветвлённые ходы в древесине[24].
У большинства же видов тело продолговатое, более или менее сплюснутое с боков, двусторонне-симметричное. Голова редуцирована, и моллюск, по сути дела, состоит из туловища и ноги[25].
Раковина |
Строение раковины двустворчатых



Раковина ископаемого двустворчатого моллюска Anadara из плиоценовых отложений Кипра с таксодонтным замком
Створки раковины у двустворчатых моллюсков чаще симметричны (например, у сердцевидок, беззубок). Однако у некоторых видов может наблюдаться асимметрия створок. Так, у устрицы створка, на которой животное лежит, выпуклая, а другая створка плоская и играет роль крышечки, прикрывая первую. Подобное явление имеет место и в случае лежащего на дне гребешка. Очень отчётливо выражена асимметрия створок у моллюсков ископаемой группы рудистов: одна из створок, погружённая в грунт, конусовидная, а другая служила крышечкой[21].
На одной из створок раковины ближе к лигаменту заметен округлый шершавый выступ — верхушка (макушка) створки раковины (см. рисунок). Это самая старая часть раковины, так как раковина прирастает по краю с противоположной стороны. Край створки, несущий макушку, называется спинным, или верхним, или замочным, краем, противоположный ему край — брюшным, или нижним, краем[26]. На переднем конце раковины располагаются нога и биссусовая железа (если есть), а на заднем конце открываются отверстия сифонов. Различают также передний и задний края створки. На заднем крае располагается мантийный синус (см. раздел Мантия и мантийная полость), отпечаток заднего мускула-замыкателя больше (см. раздел Мускулатура)[26]. У некоторых двустворчатых концы верхнего края, оттянутые от макушки вперёд и назад, образуют ушки[26]. Если расположить раковину верхушкой раковины вверх и передним концом от себя, то створку, находящуюся слева от плоскости смыкания створок, называют левой, а створку, находящуюся справа, — правой[26][27].
Створки раковины соединены лигаментом — связкой, состоящей из утолщённого рогового слоя раковины. Этот факт свидетельствует, что раковина двустворчатых произошла от цельной. У двустворчатого моллюска Arca между макушкой и верхним краем располагается связочная площадка (арея). На ней имеются бороздки (шевроны), представляющие собой следы прикрепления связки, передвигающейся по мере роста раковины[26].
Стенка раковины состоит из трёх слоёв: наружного конхиолинового (периостракум), внутреннего известкового (остракум) и нижнего перламутрового (гипостракум). Минеральный компонент раковины (то есть входящий в состав остракума и гипостракума) может быть представлен исключительно кальцитом, как у устриц, или кальцитом и арагонитом. Иногда арагонит формирует также перламутровый слой, как в случае отряда Pterioida. У остальных моллюсков слои арагонита и кальцита чередуются[28]. Лигамент и биссус, если они обызвествляются, состоят из арагонита[28]. Периостракум, состоящий из твёрдого органического вещества (конхиолина), представляет собой наружный слой и секретируется краем мантии. На вершинах створок конхиолиновый слой нередко стирается. Этот верхний слой имеет покровительственную окраску, обычно коричневую или оливковую[29]. Перламутровый слой состоит из мелких минеральных пластиночек, соединённых конхиолином. Такое строение перламутра вызывает интерференцию света, благодаря чему он переливает всеми цветами радуги. Перламутровый слой утолщается с возрастом моллюска и приростом его раковины[21].

Образование жемчужины в перламутровом слое раковины морской жемчужницы рода Pinctada
Если между створкой раковины и мантией попадает инородная частица, она обволакивается концентрическими слоями перламутра. Так образуется жемчужина[21].
Прирост раковины происходит за счёт постепенного отложения краем мантии конхиолинового слоя, а также накопления в раковине минеральных веществ. На раковине заметны концентрические линии, свидетельствующие о её неравномерном приросте в изменяющихся условиях окружающей среды (линии прироста). Однако определение возраста раковины по количеству таких концентрических линий не является достаточно точным. Более точно о возрасте моллюска можно сказать по количеству слоёв на поперечном срезе раковины.
Как упоминалось выше, две створки раковины соединяются при помощи лигамента, который состоит из двух кератиновых белков — тензилиума и резилиума. У различных групп двустворчатых лигамент может быть внутренним и наружным. Его функцией, помимо скрепления створок, является участие в раскрытии раковины.
У большинства двустворчатых моллюсков на створках раковины расположен замо́к — система зубцов и соответствующих им углублений на спинном крае раковины с внутренней поверхности. Замок препятствует смещению створок относительно друг друга. Замок может быть равнозубый (таксодонтный) и разнозубый (гетеродонтный). У некоторых Bivalvia он редуцирован, например, у беззубки (Anodonta)[21]. Строение замка является важным для идентификации моллюсков[30].
У некоторых двустворчатых раковина в значительной мере редуцирована. Так, у корабельного червя она прикрывает лишь 1/20 тела[31].
Мантия и мантийная полость |

Вскрытая мантийная полость европейской жемчужницы (Margaritifera margaritifera). 1 — задний замыкатель, 2 — передний замыкатель, 3 — верхняя полужабра, 4 — нижняя полужабра, 5 — выводной сифон, 6 — вводной сифон, 7 — нога, 8 — замок, 9 — закругление, 10 — мантия, 11 — верхушка раковины

Венериды с высунутыми сифонами
У двустворчатых моллюсков мантия имеет форму двух складок кожи, свешивающихся со спины по бокам к брюшной стороне. Снизу её складки могут быть свободными (как у беззубки) или срастаться, оставляя лишь отверстия для ноги и сифонов. По краю мантии иногда могут развиваться маленькие щупальца и глаза[25] (подробнее см. раздел Органы чувств). Наружный слой мантии выделяет раковину, а внутренний выстлан мерцательным эпителием, биение ресничек которого обеспечивает ток воды в мантийной полости[21]. Помимо раковины, мантия образует также лигамент, биссусову железу и замок[32].
У роющих форм мантия образует сифоны — две длинные трубки, по нижней из которых (вводной сифон) вода входит в мантийную полость, а по верхней (выводной сифон) — выходит. С током воды в мантийную полость доставляются кислород и пищевые частицы[21].
Как и у всех моллюсков, у двустворчатых мантия образует мантийную полость, в которую входит мантийный комплекс органов: нога, две жабры, две ротовые лопасти и осфрадии. В мантийную полость также открываются отверстия пищеварительной, половой и выделительной систем[21].
У большинства двустворчатых моллюсков на внутренней стороне раковины заметна линия, идущая параллельно краю раковины и часто соединяющая следы от двух мускулов-аддукторов (замыкателей). Её называют паллиальной (мантийной) линией, она представляет собой линии прикрепления мантии к створке раковины. Прикрепление осуществляется узким рядом небольших мускулов-ретракторов. С помощью этих мускулов моллюск может в случае опасности спрятать внутрь раковины выступающий край мантии. Втягиваться могут и сифоны (в нормальном состоянии высунутые из раковины). Для этого служит специальное кармановидное углубление в мантийной полости. На створке раковины этому углублению соответствует паллиальный синус, или мантийный синус, или мантийная бухта, или сифонный залив, — направленное внутрь искривление паллиальной линии[26][27].
Нога |

Мидия рода Mytilus, прикрепившаяся к субстрату биссусными нитями
Нога (мускулистый непарный вырост брюшной стенки) у двустворчатых моллюсков клиновидная, служит для зарывания в грунт и ползания. У наиболее примитивных форм (отряд Protobranchia) нога, как и у брюхоногих, с плоской ползательной подошвой[25]. Некоторые двустворчатые, прикрепляющиеся к субстрату, имеют в ноге особую биссусовую железу, выделяющие биссусные нити, при помощи которых моллюск «прирастает» к поверхности дна (мидии). У многих двустворчатых, ведущих неподвижный образ жизни, нога редуцируется полностью (устрицы)[31].
Внутреннее строение |

Вскрытая мидия съедобная (Mytilus edulis)
Мускулатура |
Основными мышцами в теле пластинчатожаберных моллюсков являются передний и задний мускулы-замыкатели (аддукторы), хотя передний замыкатель у некоторых видов может быть редуцирован или полностью утрачен. Сокращаясь, эти сильные мускулы закрывают створки, а при их расслаблении створки открываются. Кроме того, в механизме раскрывания створок задействован лигамент. Когда раковина закрыта, он, как пружина, находится в натянутом состоянии. При расслаблении замыкателей он возвращается в исходное положение, открывая створки[31]. У двустворчатых моллюсков, лежащих на одной створке (например, устриц и мидий), передний аддуктор утрачен, а задний занимает центральное положение. Bivalvia семейства лим (Limidae), плавающие за счёт хлопания створками, тоже имеют единственный центральный замыкатель. Замыкатели состоят из двух типов мышечных волокон: поперечно-полосатых, предназначенных для быстрых движений, и гладких, поддерживающих длительное напряжение мускула[29].
Как упоминалось выше, мантия прикрепляется к раковине за счёт мелких мышц, образующих дугообразный след на створке раковины — паллиальную линию. Парные протракторные (сгибательные) и ретракторные (разгибательные) мускулы обеспечивают движения ноги двустворчатого. У Bivalvia, лишённых ноги, этих мускулов нет. Другие парные мышцы контролируют сифоны и биссусову железу[27][29].
Пищеварительная система |
В связи с пассивным способом питания путём фильтрации пищеварительная система двустворчатых имеет некоторые особенности. Вода, поступающая через вводной сифон, направляется к переднему концу тела, омывая жабры и 2 пары длинных треугольных ротовых лопастей. На жабрах и ротовых лопастях имеются чувствительные клетки (органы вкуса) и мелкие желобки, по которым пищевые частицы транспортируются в рот, находящийся около переднего замыкателя. Изо рта пища поступает в короткий пищевод и затем в мешковидный энтодермальный желудок. Так как у двустворчатых редуцирована голова, то глотка, радула и слюнные железы у них отсутствуют. В желудок открывается несколько пищеварительных желёз, часто через пару дивертикулов, как, например, двухлопастная печень. Печень не только выделяет пищеварительные ферменты: её клетки также фагоцитируют пищевые частицы. Таким образом, у двустворчатых имеется внутриклеточное пищеварение. Кроме того, в желудке есть кристаллический стебелёк, состоящий из мукопротеинов и ферментов (амилазы, гликогеназы и др.). Стебелёк располагается в особом слепом мешковидном выросте[33] и вдаётся в просвет желудка. Располагающиеся там реснички заставляют стебелёк вращаться, отделяя ферменты и перемешивая содержимое желудка. Благодаря постоянному движению пищевых частиц в желудке возможна их сортировка в его заднем конце: мелкие частицы отправляются в пищеварительные железы и там поглощаются путём фагоцитоза, а более крупные направляются в кишечник. От желудка отходит средняя кишка, которая далее делает несколько изгибов и направляется по спинной стороне туловища к заднему концу, переходит в заднюю кишку, которая открывается анальным отверстием в мантийную полость над задним замыкателем[34]. Экскременты с током воды выбрасываются через выводной сифон наружу[35]. Задняя кишка обычно проходит через желудочек сердца (специфическая особенность двустворчатых)[36][37].
Питание и пищеварение у двустворчатых синхронизированы с суточными и приливно-отливными ритмами[38].
Описанные выше особенности пищеварительного тракта характерны для фильтрующих двустворчатых. У хищных двустворчатых стебелёк может быть сильно редуцирован, но зато в ряде случаев имеется мускульный желудок, выстланный хитином, в котором пища перетирается ещё до начала пищеварения. В других случаях пищеварительный тракт хищных двустворчатых схож с таковым у двустворчатых-фильтраторов[39].
Нервная система |
Как и у большинства моллюсков, нервная система у двустворчатых разбросанно-узлового типа. У них она устроена проще, чем у брюхоногих. Из-за редукции головы церебральные ганглии слились с плевральными; так образовались парные цереброплевральные сдвоенные узлы, располагающиеся по обе стороны от пищевода и соединяющиеся над глоткой тонкой церебральной комиссурой. Образование цереброплевральных узлов путём срастания ганглиев доказывается тем, что у примитивных Protobranchia плевральные узлы ещё обособлены от церебральных. Они иннервируют мантийную полость и органы чувств (кроме осфрадиев). В ноге имеются педальные ганглии, иннервирующие ногу, связанные коннективами с цереброплевральными узлами. Под задним мускулом-замыкателем находится третья пара узлов — висцеропариетальные, контролирующие внутренние органы, жабры и осфрадии[40]. Они связаны ещё более длинными коннективами с цереброплевральными узлами[37]. Особенно хорошо третья пара узлов развита у плавающих двустворчатых. Двустворчатые с длинными сифонами могут иметь специальные сифональные ганглии, контролирующие сифоны[41].
Органы чувств |

Глазки морского гребешка, расположенные по краю мантии
Органы чувств у двустворчатых моллюсков развиты слабо. В ноге имеются статоцисты — органы равновесия, иннервирующиеся церебральными ганглиями. В мантийной полости у основания жабр находятся осфрадии — органы химического чувства; возможно, осфрадии двустворчатых негомологичны осфрадиям брюхоногих[42]. Отдельные рецепторные клетки разбросаны на жабрах, ротовых лопастях, по краю мантии и на сифонах[43]. Осязательную функцию выполняют также щупальца, развивающиеся по краю мантии[44]. У хищных Bivalvia из отряда Anomalodesmata сифоны окружены щупальцами, чувствительными к вибрациям; с их помощью моллюски обнаруживают добычу[45].
Многие двустворчатые лишены глаз, однако представители групп Arcoidea, Limopsoidea, Mytiloidea, Anomioidea, Ostreoidea и Limoidea имеют простые глазки, расположенные по краю мантии. Они состоят из ямки, выстланной светочувствительными клетками, и светопреломляющего хрусталика[46]. У морских гребешков имеются инвертированные глазки довольно сложного строения[37], состоящие из хрусталика, двухслойной сетчатки и вогнутой светоотражающей поверхности[47]. Известны также случаи образования глаз на сифонах у сердцевидок[43]. У всех двустворчатых есть светочувствительные клетки, благодаря которым моллюск определяет, когда его полностью накроет тень[41].
Дыхательная система |
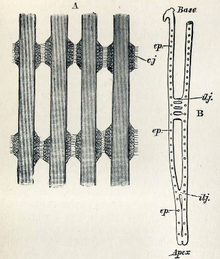
Жабры мидии съедобной (Mytilus edulis). А — участок четырёх жаберных нитей; соседние нити соединяются друг с другом при помощи ресничек (cj). B — строение жаберной нити
Дыхательная система представлена жабрами (ктенидиями). Особенности строения жабр варьируют у различных групп двустворчатых.
Первичножаберные (Protobranchia), являющиеся наиболее примитивными двустворчатыми, имеют пару типичных ктенидиев с жаберными лепестками.- У нитежаберных (Filibranchia) имеются нитевидные жабры. Нитевидные жабры характеризуются тем, что их жаберные лепестки удлинились в нити, образующие сначала нисходящее, а затем восходящее колено. Соседние нити скрепляются друг с другом при помощи жёстких ресничек, формируя пластинки; у части представителей жаберные нити свободны[48]. Нитевидные жабры свойственны мидиям, устрицам, гребешкам.
- У отряда Eulamellibranchia имеются пластинчатые жабры. Это дальнейшая модификация нитевидных жабр: в них возникают перегородки между соседними нитями, а также восходящими и нисходящими участками одной нити. Так образуются жаберные пластинки. Каждая жабра состоит при этом из двух полужабер: наружной, прилегающей к мантии, и внутренней, прилегающей к ноге. Таким образом, у Eulamellibranchia имеется 4 жабры, но каждая из них соответствует лишь одной половине настоящего ктенидия[48]. Такие жабры есть у перловиц, беззубок.
- У перегородчатожаберных (Septibranchia) жабры редуцированы и преобразованы в жаберную перегородку с порами. Перегородка отгораживает верхнюю часть мантийной полости, образуя дыхательную полость. Её стенки пронизаны кровеносными сосудами, где и происходит газообмен[49].
Наконец, у видов, лишённых жабер (как, например, у представителей подкласса Anomalodesmata), газообмен идёт через стенку мантийной полости[50].
Двустворчатые, живущие в приливно-отливной зоне, способны несколько часов выживать без воды, плотно закрыв створки. Некоторые пресноводные формы, будучи вытащенными на воздух, слегка приоткрывают створки, так что у них происходит газообмен с атмосферным воздухом[50].
Кровеносная система |

Двустворчатый моллюск Arca zebra, имеющий два сердца
Кровеносная система двустворчатых, как и всех моллюсков, незамкнутая, то есть кровь циркулирует не только по сосудам, но и по лакунам (промежуткам между органами). Сердце расположено на спинной стороне и состоит из 1 желудочка и 2 предсердий. Как упоминалось выше, сквозь желудочек проходит задняя кишка. Этот факт объясняют тем, что сердце закладывается в эмбриогенезе как парное по бокам кишки, а потом эти зачатки соединяются над и под кишкой (парное происхождение сердца у двустворчатых подтверждается наличием двух сердец у представителей рода Arca)[51]. У примитивных форм отряда Protobranchia слияние происходит только над кишкой[52].
От желудочка отходят мощные передняя и задняя аорты, ветвящиеся в артерии; из них кровь (гемолимфа) изливается в лакуны и отдаёт кислород тканям. Передняя артерия идёт вперёд над кишкой и снабжает кровью внутренности, ногу и переднюю часть мантии, а задняя направлена назад под кишечником и вскоре распадается на задние мантийные артерии[52]. У некоторых двустворчатых моллюсков имеется лишь одна аорта[53]. Далее уже ставшая венозной кровь собирается в крупную продольную лакуну под сердцем и направляется в приносящие жаберные сосуды. Потом насыщенная кислородом артериальная кровь по выносящим сосудам возвращается из жабр в сердце. В выносящие жаберные сосуды изливается также кровь, которая, минуя жабры, проходит через почки, где освобождается от продуктов обмена[51].
Кровь двустворчатых обычно лишена какого-либо дыхательного пигмента, хотя у представителей семейств Arcidae и Limidae имеется гемоглобин, растворённый непосредственно в плазме крови[39]. У хищного двустворчатого моллюска Poromya в крови есть красные амёбоциты, содержащие гемоглобин[54].
Выделительная система |
Выделительная система двустворчатых, как и у большинства моллюсков, представлена парными нефридиями (почками). Почки двустворчатых с железистыми стенками называют боянусовыми органами. Почки представляют собой длинные V-образные трубки, которые одним концом открываются в перикард, другим — в мантийную полость, откуда продукты обмена уносятся с током воды[55][51]. По своему происхождению они являются типичными целомодуктами[52].
Кроме почек, выделительную функцию также выполняет стенка перикарда, видоизменённая в парные перикардиальные железы. Иногда они обособляются от остального перикарда в виде двух мешковидных образований — кеберовых органов[51]. Продукты выделения этих желёз попадают в перикард, а оттуда через почки транспортируются наружу[52].
Осморегуляция |
Двустворчатые относятся к пойкилосмотическим животным, то есть они не способны сохранять более или менее постоянное осмотическое давление полостных и тканевых жидкостей при изменении солёности воды. Они также не могут поддерживать осмотическое давление ниже, чем во внешней среде[56].
У пресноводных двустворчатых, как и у пресноводных брюхоногих, через почки выделяется много воды; в почках же реабсорбируются соли, поэтому по сравнению с кровью (гемолимфой) моча пресноводных Bivalvia сильно гипоосмотична[57].
Эпителий, выстилающий мантию и жабры двустворчатых, обладает способностью поглощать соли из воды, которая проходит через мантийную полость[57].
Половая система |

Створки раковины Arca noae
Двустворчатые моллюски раздельнополы, однако имеются и случаи гермафродитизма (так, у вида Arca noae был установлен протандрический гермафродитизм, при котором сначала особи функционируют как самцы, потом как самки[58]). У некоторых видов, например Thecaliacon camerata, резко выражен половой диморфизм[33]. Гонады и протоки (семяпроводы и яйцеводы) парные; гонады залегают в передней части тела, близко к кишке, заходя в основание ноги, и имеют вид двух дольчатых гроздевидных образований. Однако у некоторых видов половые протоки отсутствуют, и гаметы выходят из гонад через разрывы тканей в мантийную полость[51]. У примитивных Protobranchia, а также ряда других двустворчатых (Pecten, Ostrea и др.) гонады открываются в почки[59].
У некоторых видов, например представителей рода Lasaea, мужские половые клетки выходят наружу через сифон, а далее с током воды затягиваются в мантийную полость самок, где и происходит оплодотворение[60]. Потомство у таких видов развивается в мантийной полости матери и покидает её на стадии личинки — велигера или молодой особи[61]. У большинства же видов оплодотворение наружное[59][33]. В этом случае самки и самцы выбрасывают сперму и яйцеклетки в толщу воды. Этот процесс может идти непрерывно или запускаться факторами внешней среды, например продолжительностью дня, температурой воды и наличием спермы в воде. Некоторые пластинчатожаберные выпускают гаметы понемногу, другие же — крупными частями или все одновременно. Массовый выброс гамет иногда происходит, когда все двустворчатые в округе синхронизированно высвобождают половые клетки[62].
Отличия от плеченогих |

Строение раковины брахиопод. Тёмно-серым отмечена ножная створка раковины, светло-серым — плечевая, розовым — ножка
Плеченогие (Brachiopoda) — морские организмы, напоминающие двустворчатых моллюсков раковиной из двух створок и близким размером. Однако они не имеют близкого родства с моллюсками, и их выделяют в отдельный тип. Сходство их внешнего строения является конвергентным, то есть обусловлено схожим образом жизни. В настоящее время брахиоподы менее распространены, чем двустворчатые[63].
Обе эти группы животных имеют раковину из двух створок, однако строение раковины у них коренным образом отличается. У плеченогих створки раковины располагаются на дорсальной (спинной) и вентральной (брюшной) сторонах тела, а у двустворчатых они прикрывают левую и правую сторону тела и обычно являются зеркальными копиями друг друга. Кроме этого, у плеченогих имеется специфический орган для фильтрации — лофофор, отсутствующий у двустворчатых. Наконец, раковины брахиопод могут состоять как из фосфата кальция, так и карбоната кальция, а у двустворчатых в их состав входит только карбонат кальция[64].
Жизненный цикл |
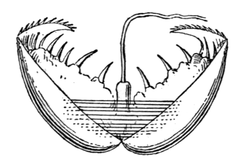
Глохидии

У двустворчатых, как и у всех моллюсков, спиральное дробление. Оно идёт примерно так же, как и у брюхоногих. Большинство двустворчатых развиваются с метаморфозом. Обычно из оплодотворённых яиц выходит планктонная личинка — велигер (парусник). Образованию велигера предшествует стадия трохофоры, проходящая в яйце. Образование трохофоры происходит довольно быстро и занимает несколько часов или дней. На спинной стороне трохофоры закладывается раковина в виде цельной пластинки, которая лишь позднее перегибается по срединной линии, становясь двустворчатой, причём место перегиба сохраняется в виде лигамента. Верхняя часть трохофоры с венчиком ресничек становится парусом велигера — покрытым длинными ресничками диском, служащим для плавания. Двустворчатая раковина покрывает всё тело велигера, при плавании парус выставляется из раковины. Организация велигера очень близка к организации взрослого моллюска: он имеет зачаток ноги, мантию, ганглии, желудок, печень и другие органы, однако органами выделения остаются протонефридии[59]. В дальнейшем велигер оседает на дно, закрепляется биссусовой нитью, теряет парус и превращается во взрослого моллюска[60].
У некоторых пресноводных моллюсков (перловиц, беззубок) имеется особая личинка — глохидий, имеющая тонкостенную двустворчатую раковину с округлыми створками и крючками на брюшном крае. Большинство органов глохидия ещё недоразвито: отсутствуют жабры, нога рудиментарна[59]. У таких моллюсков оплодотворение происходит в мантийной полости самки, и глохидии развиваются в её жабрах. Глохидий способен плавать, хлопая створками. С помощью биссусовой нити глохидий прикрепляется к жабрам проплывающих мимо рыб и с помощью зубцов внедряется в их ткани, где и происходит его дальнейшее развитие. Позднее маленький моллюск, развившийся из глохидия, выпадает на дно через разрыв опухолевых бугорков на коже рыб. Таким образом, глохидий является эктопаразитом, а рыбы обеспечивают его распространение[60]. Тем не менее, серьёзного ущерба рыбе глохидии не приносят[65].
У некоторых пресноводных двустворчатых моллюсков развитие прямое. Так, у шаровок (Sphaerium) яйца развиваются в мантийной полости, и из них вылупляются сформированные маленькие моллюски[60].
У большинства двустворчатых личинки, вышедшие в воду, питаются диатомовыми водорослями и другим фитопланктоном. В умеренных регионах около 25 % видов являются лецитотрофными, то есть ещё не ставшие взрослыми особи зависят от питательных веществ, запасённых в желтке яйца, из которых основным источником энергии являются липиды. Чем больше период до первого самостоятельного питания личинки, тем крупнее яйцо и больше в нём запас желтка. Энергозатраты на образование таких яиц велики, и обычно такие богатые желтком яйца формируются в небольших количествах. Так, балтийская макома (Macoma balthica) образует немного яиц с большим запасом желтка. Выходящая из яйца личинка живёт за счёт запасов энергии яйца и не питается. Приблизительно через 4 дня они переходят в D-стадию личинки, в которой у них формируется первая раковина из D-образных створок. Такие личинки имеют невысокий потенциал для расселения. Мидия съедобная (Mytilus edulis) образует чрезвычайно много яиц, бедных желтком, так что образующиеся из них личинки должны рано начинать питаться, чтобы выжить и расти. Они могут распространяться более широко, так как дольше ведут планктонный образ жизни[66].
У некоторых пресноводных двустворчатых семейства унионид (Unionidae) стратегия размножения весьма необычна. Мантия самки выдаётся из раковины и имитирует маленькую рыбу: на ней появляются отметины, напоминающие чешую рыб, и ложные глаза. Эта приманка привлекает внимание настоящих рыб. Некоторые рыбы рассматривают её как добычу, другие видят в ней особь своего вида. Рыбы подплывают ближе, чтобы рассмотреть приманку, и тогда моллюск выбрасывает колоссальное количество глохидиев на рыбу, которые внедряются в её жабры и покровы[65].
Рассмотренные выше случаи вынашивания молоди в жабрах в мантийной полости, а также в выводковых камерах (Mineria minima) можно рассматривать как проявление заботы о потомстве[33].

Arctica islandica — одно из самых долгоживущих животных, максимальный возраст — 507 лет[67]
К двустворчатым относятся самые долгоживущие — достигающие возраста свыше 400 лет — представители царства животных. Этому классу животных принадлежит рекорд по количеству видов с продолжительностью жизни более 150 лет. Так, у морского моллюска Panopea abrupta, пресноводного моллюска Margaritifera margaritifera и океанского венуса Arctica islandica максимальная продолжительность жизни составляет 163 года, 190[68] и 507 лет соответственно[67].
Для двустворчатых моллюсков была установлена связь между максимальным размером раковины, скоростью роста, возрастом полового созревания и продолжительностью жизни, а именно:
- максимальный размер раковины положительно связан с наибольшей продолжительностью жизни;
- срок жизни после полового созревания прямо пропорционален возрасту полового созревания;
- чем раньше двустворчатый моллюск достигает своих максимальных размеров, тем меньше продолжительность его жизни[68].
Так, средний возраст полового созревания уже упоминавшегося долгоживущего моллюска Arctica islandica составляет около 10 лет, при этом массы в 20 г он достигает за 25—30 лет. В то же время двустворчатый моллюск Spisula solidissima достигает той же массы за 2—3 года, и иногда начинает размножаться в первый год жизни[69].
Питание |
Большинство двустворчатых моллюсков являются фильтраторами и питаются, в частности, фитопланктоном. У наиболее примитивных двустворчатых — Protobranchia — встречаются разные модели питания, в том числе соскабливание детрита с морского дна; возможно, эта модель питания была исходной для двустворчатых, пока они ещё не приобрели приспособления к питанию путём фильтрования. Они ощупывают субстрат с помощью пары щупалец, образуемых краем мантии. Щупальца покрыты слизью и снабжены ресничками. Щупальца собирают со дна пищевые частицы и сортируют их, отбрасывая слишком крупные и направляя остальные в рот[39]. Особенности пищеварительной системы двустворчатых, связанные с фильтрующим типом питания, рассматривались выше.

Хищный двустворчатый моллюск Poromya granulata
Небольшое число Bivalvia, как, например, Poromya granulata, являются хищниками. Они относятся к надотряду перегородчатожаберные[70]. Специальные мускулы направляют воду во вводной сифон, преобразованный в капюшоновидный орган, который засасывает мелких ракообразных и червей. Сифон может быстро сокращаться и вворачивается в специальную камеру, а добыча передается на ротовые лопасти, которые доставляют её ко рту [1]. Пищеварительный тракт хищных пластинчатожаберных модифицирован таким образом, что в нём могут перевариваться крупные пищевые частицы[54].
Весьма необычный представитель двустворчатых Entovalva nhatrangensis является эндосимбионтом и встречается только в пищеводе голотурий. Складки мантии у этого вида целиком прикрывают маленькую раковину. Когда голотурия зарывается в донный ил, моллюск пропускает воду через свои жабры и фильтрует органические частицы. Чтобы его самого не унесло потоком воды, моллюск крепко прикрепляется биссусом к пищеводу хозяина. Голотурии при этом никакого вреда не наносится[71].

Корабельный червь рода Teredo
Некоторые двустворчатые моллюски вступают в симбиоз с бактериями, участвующими в их процессе пищеварения, или же питаются за их счёт. Например, у некоторых солемий (Solemya), обитающих в среде, богатой сероводородом (побережья, занятые свалками, промышленными предприятиями), пищеварительная система атрофируется в разной степени, вплоть до полной редукции. Питаются же такие солемии предположительно за счёт окисляющих сероводород бактерий, поселяющихся в жабрах моллюска либо свободно живущих в грунте, но которых моллюск отфильтровывает и переваривает в клетках жабр[72]. Представители семейства корабельные черви (Teredinidae) имеют червеобразную форму тела и питаются древесиной деревянных подводных предметов, в которую вгрызаются сверлильным аппаратом раковинки (которая редуцируется до двух маленьких пластинок на переднем конце тела). В результате древесина, пронизанная многочисленными ходами от корабельного червя, становится похожей на губку и легко разрушается. Переваривание древесины у корабельных червей осуществляют симбиотические бактерии[73].
Образ жизни |

Морской финик Lithophaga truncata
Большинство двустворчатых ведут сидячий (прикрепленный) или малоподвижный образ жизни, проводя всю жизнь на том месте, где они осели на стадии личинки. Все свободноживущие двустворчатые (кроме сверлящих форм) является бентосными организмами, они живут на поверхности дна или зарываются в субстрат: песок, ил, гальку, фрагменты кораллов. Многие из них живут в приливно-отливной (литоральной) зоне, где грунт остаётся влажным даже в отлив. Зарываясь в грунт, двустворчатые предохраняют себя от ударов волн, обезвоживания и перегрева во время малой воды, а также от перепадов солёности воды, вызванных дождями. Кроме того, это спасает от многих хищников[74]. Их главная стратегия такова: во время прилива двустворчатые выставляют свои сифоны для питания и дыхания, а в отлив зарываются на большую глубину или плотно закрывают створки раковины[74]. Для зарывания в грунт они используют свою мускулистую ногу. Механизм зарывания состоит в следующем. Сначала моллюск расслабляет мускулы-замыкатели и широко раскрывает створки раковины, заякориваясь в определённой позиции. В это время нога погружается в грунт. Потом он растягивает конец ноги, сокращает замыкатели и закрывает раковину, сокращает ногу и уходит глубже в грунт. Повторяя эти движения, моллюск может закапываться на большую глубину[75].
Другие формы, как, например, мидии, прикрепляются к твёрдым субстратам при помощи биссуса — нити, состоящей из кератиновых белков. Некоторые двустворчатые прочно прирастают к субстрату створкой раковины (Ostrea, Pinctada)[76]. Такие двустворчатые более подвержены нападениям хищников, чем зарывающиеся. Некоторые хищные брюхоногие, например трубачи (Buccinidae) и иглянки (Muricidae), питаются двустворчатыми, просверливая их раковины. Морская улитка Nucella lamellosa пробуравливает радулой ходы в раковинах двустворчатых моллюсков (обычно мидии съедобной) и выделяет при этом разлагающий раковину секрет. Потом она вставляет в образовавшееся отверстие растяжимый хоботок и высасывает тело жертвы. Чтобы просверлить раковину двустворчатого моллюска, улитке нужно всего несколько часов, так что в этом отношении двустворчатые, живущие в зоне литорали, имеют преимущество, поскольку могут подвергаться нападению улиток только во время прилива[77].
Некоторые двустворчатые, в том числе устрицы семейств хамиды (Chamidae), Anomiidae, спондилиды (Spondylidae), Plicatulidae приклеиваются к камням, скалам и более крупным твёрдым раковинам[78]. Иногда устрицы образуют густые скопления в неритической зоне и, как и большинство двустворчатых, являются фильтраторами[30].

Шаровка роговая (Sphaerium corneum) способна карабкаться по водным растениям с помощью длинной гибкой ноги
Хотя многие несидячие двустворчатые используют ногу для небольших перемещений вокруг или для закапывания, представители семейства Sphaeriidae весьма проворно карабкаются по водным растениям с помощью своей длинной и гибкой ноги. Так, шаровка роговая (Sphaerium corneum) лазает по растениям элодеи по берегам озёр и прудов; так моллюск находит лучшую позицию для фильтрования[79].
Как говорилось выше, морские гребешки способны к реактивному движению, хлопая створками раковины за счёт мощного мускула-замыкателя; таким образом гребешки могут переплывать на небольшие расстояния[80][81].
Камнеточцы из семейства морских фиников (Lithophagidae) при помощи специального кислого секрета мантийной железы протачивают ходы в известняке и закрепляются в них при помощи биссуса, выставляя наружу сифоны[80]. Известно, что колонны храма Сераписа близ Неаполя проточены Litophaga до высоты человеческого роста[76]. Камнеточцы, способные повреждать известняки, песчаники и даже бетон, также образуют семейство камнеточцев фолад (Pholadidae). Кроме того, камнеточец Pholas способен к биолюминесценции. Именно на этом моллюске в 1887 году Дюбуа открыл люциферин-люциферазную реакцию. Свечение испускает край мантии; также Pholas может выделять люминесцирующую слизь[82].
Весьма специфический образ жизни ведут представители семейства древоточцев, или корабельных червей (Teredinidae)[70].
Среди двустворчатых моллюсков есть и эндосимбионты, например, уже упоминавшийся эндосимбионт голотурий Entovalva nhatrangensis.
Экология |
Роль в экосистемах |
Особо велика роль двустворчатых моллюсков в биологической очистке вод. Двустворчатые моллюски выступают как биофильтраторы, очищающие водоёмы от органического загрязнения. Кроме того, они поглощают и накапливают в своём теле тяжёлые металлы, тем самым способствуя очищению водоёмов от химического загрязнения. Их фильтрующая активность довольно велика — в среднем 1 л воды в час[83]. Выяснено, что мидии, заселяющие 1 м² дна, за сутки могут профильтровать 280 м³ воды[76].
Кроме того, замечено, что в районах разведения моллюсков повышается общая продуктивность океана, в том числе и численность рыб, которые питаются моллюсками и прочими беспозвоночными, обитающими в их скоплениях[83].
Двустворчатые моллюски играют большую роль в образовании осадочных известковых пород. Из раковин двустворчатых немалой частью состоят такие горные породы, как мрамор, известняки, ракушечник. Стоит отметить, что наиболее многочисленные виды ископаемых Bivalvia являются руководящими формами для определения возраста пластов Земли[84].
Взаимоотношения с другими организмами |
Естественные враги |

Limaria fragilis. Моллюск имеет белую раковину с тонкими асимметричными створками и красную мантию со щупальцами по краю[85]
Прочная раковина и её округлая форма делают двустворчатых моллюсков малодоступными для хищников. Тем не менее многие животные питаются ими. Среди них есть и рыбы, например обыкновенный карп (Cyprinus carpio)[86]; среди птиц двустворчатыми питаются, например, кулик-сорока (Haematopus ostralegus), раскусывающий раковину специально адаптированным клювом[87], и серебристая чайка (Larus argentatus), разбивающая раковины, сбрасывая на них камни[88]. Калан (Enhydra lutris) питается многими двустворчатыми моллюсками, разбивая их раковины камнями[89]. Морж (Odobenus rosmarus) является одним из главных хищников арктических вод, питающихся двустворчатыми[90]. Из беспозвоночных естественными врагами двустворчатых моллюсков являются крабы[78],
морские звёзды и осьминоги[91]. Некоторые брюхоногие моллюски: устричное сверло (Urosalpinx), Eupleura caudata, Polynices heros, Neverita duplicata — тоже уничтожают двустворчатых, в том числе ценных промысловых животных — устриц и мидий[92].
Механизмы защиты от врагов у двустворчатых моллюсков разнообразны. Некоторые закапываются в грунт (как, например, Siliqua patula, способная закопаться за 7 секунд[93]); как упоминалось выше, морские гребешки и некоторые другие моллюски способны плавать, хлопая створками раковины. Другие пластинчатожаберные способны ускакать от угрозы на ноге, используя её как пружину[94]. Двустворчатые, имеющие сифоны, могут скрыться внутри раковины, выставив сифоны наружу; если хищник оторвёт их, они регенерируют[95]. Лимы, как, например, Limaria fragilis, в случае раздражения выделяют токсичные вещества[96].
В 2016 году у двустворчатых моллюсков был описан заразный рак, передающийся посредством морской воды и заражающий моллюсков разных видов[97].
Симбионты |

Створка раковины Corculum cardissa
Некоторые двустворчатые моллюски вступают в симбиотические взаимоотношения с протистами (а именно одноклеточными водорослями). В частности, двустворчатый моллюск Corculum cardissa вступает в симбиоз с Symbiodinium corculorum из группы динофлагеллят. Клетки протистов обнаруживаются при световой или трансмиссионной электроскопии преимущественно в тканях мантии и жабр моллюска. Протисты (зооксантеллы) поселяются также в утолщённом крае мантии гигантской тридакны (Tridacna gigas), но, в отличие от C. cardissa, в клетках жабр они не обнаружены[98][99].
Эволюция |

Раковина ископаемого плеченогого. Виден лофофор
Двустворчатые, как и большинство других обладающих скелетом групп беспозвоночных, появились в палеонтологической летописи в ходе кембрийского взрыва (около 540 млн лет назад)[100]. К этому же времени относятся первые ископаемые останки и многих других типов животных.
Раннекембрийские ископаемые моллюски Fordilla и Pojetaia считаются ранними двустворчатыми[101][102][103][104]. Кроме этих двух родов, к кембрийским двустворчатым предположительно относятся Tuarangia, Camya и Arhouriella, а также, возможно, Buluniella. Существует также версия, что двустворчатые моллюски произошли от Rostroconchia — вымершего класса моллюсков.
Иногда окаменелости образуются, когда грунт, в котором находится раковина, окаменевает вместе с ней. Иногда они образуются другими путями. Нередко окаменелостью является не сама раковина, а след, оставленный ей на камне.
В раннем ордовике произошёл большой всплеск разнообразия двустворчатых; в это время обособились группы беззубых, равнозубых и разнозубых двустворчатых. В этом периоде появляются двустворчатые, напоминающие современных (443—488 млн. лет назад)[105]. В раннем силуре жабры адаптируются к фильтрующему питанию, в течение девона и карбона появляются сифоны, а также мускулистая нога, позволяющая моллюскам закапываться в грунт[106].
В раннем палеозое (около 400 млн лет назад) господствующей группой фильтраторов в океанах были брахиоподы. К этому периоду относится около 12 тысяч их ископаемых видов[107]. Ко времени массового пермского вымирания двустворчатые достигли большого разнообразия. Этот удар нанёс большой вред двустворчатым, однако они смогли выжить в течение последующего периода — триаса. В то же время брахиоподы потеряли 95 % их видового разнообразия[64]. Некоторые учёные считают, что способность двустворчатых зарываться в грунт для спасения от хищников являлась ключевым фактором их успеха. Новые адаптации, появившиеся в различных семействах, позволили двустворчатым освоить новые экологические ниши. К таким приспособлениям относят способность к плаванию, хищнический образ жизни и другие[106].
Долго считалось, что двустворчатые моллюски гораздо лучше приспособлены к жизни в воде, чем брахиоподы, в связи с чем брахиоподы не выдержали конкуренции и занимают в последнее время лишь малые экологические ниши. Эти две группы животных описываются в учебниках как пример двух таксонов, один из которых оказался конкурентоспособнее другого и вытеснил его. Основанием для такого вывода считали факт, что двустворчатому моллюску для существования нужно меньше пищи из-за её системы открывания-закрывания раковины с энергетически эффективным лигаментом. Однако все эти доводы сейчас опровергнуты, а превосходство двустворчатых над брахиоподами объясняется, как писалось выше, более успешным прохождением первыми массового вымирания[108].
Классификация |
За прошедшие два века учёные не пришли к единому мнению относительно филогении двустворчатых моллюсков и их классификации. В ранних таксономических системах учёные классифицировали двустворчатых по одному признаку, как то: морфология раковины, строение замка или жабр. Однако в таких системах, основанных на строении одного органа, возникали проблемы в наименовании таксонов. Одна из наиболее широко распространённых систем была разработана Норманом Ньюэллом в издании Treatise on Invertebrate Paleontology, Part N[109]. Эта система была основана как на общей морфологии раковины, так и её микроструктуре, а также строении замка[110]. Поскольку эти признаки мало изменялись на протяжении существования таксона, они могут быть использованы для выделения наиболее крупных групп.
Проводимые с 2000 года исследования в области таксономии с применением кладистического анализа многих систем органов, морфологии раковин (в том числе и у ископаемых видов) и современных методов молекулярной филогенетики завершились созданием наиболее вероятной схемы филогенетических взаимоотношений двустворчатых[111][112][113][114][115]. Основываясь на результатах этих исследований, в 2010 году Билер, Картер и Коэн (англ. Bieler, Carter & Coan) предложили новую классификацию двустворчатых. В 2012 году эта классификация была доработана специалистами World Register of Marine Species (WoRMS). Сейчас продолжаются молекулярно-филогенетические исследования по выявлению наиболее родственных групп двустворчатых и их группировке в соответствующие таксоны[116][117].
Ниже приведён обзор некоторых систем классификации двустворчатых, разработанных в разное время.
Практическая классификация Р. К. Мура |
Р. К. Мур в 1952 году разработал практическую классификацию двустворчатых на основании строения раковины, жабр и зубов замка. Она выглядит следующим образом:
- Подкласс Prionodesmacea
- Отряды
- Paleoconcha
Taxodonta — Гребенчатозубые: много мелких зубцов;
Schizodonta — Расщеплённозубые: крупные раздваивающиеся зубцы;
Isodonta — Равнозубые: равные зубцы;
Dysodonta — Связочнозубые: нет зубцов, створки удерживаются только лигаментом.
- Подкласс Teleodesmacea
- Отряды
Heterodonta — Разнозубые: зубцы различны;
Pachydonta: крупные различающиеся деформированные зубцы (в том числе рудисты);
Desmodonta: зубцы отсутствуют или неправильной формы; основную роль в скреплении створок играет лигамент.
У Prionodesmacea призматическая структура раковины, в раковинах есть перламутровый слой; не срослись складки мантии, плохо развиты сифоны; зубцы отсутствуют или неспециализированы. Жабры — от простых ктенидиев (Protobranchia) до пластинчатых (Eulamellibranchia). У представителей Teleodesmacea, напротив, раковина фарфоровидная, лишённая перламутра. Складки мантии, как правило, срастаются, сифоны хорошо развиты, зубцы замка специализированы. У большинства пластинчатые жабры.
Классификация 1935 года |
В 1935 году Иоганнес Тиле в своей работе Handbuch der systematischen Weichtierkunde («Руководство по систематической малакологии») представил классификацию двустворчатых моллюсков, основанную на работе Коссманна и Пейрота 1909 года. Согласно системе Тиле, двустворчатые подразделяются на три отряда. К Taxodonta относятся виды с таксодонтным замком, состоящим из нескольких параллельных рядов мелких зубцов, располагающихся перпендикулярно краю замка. К Anisomyaria принадлежат виды, у которых или один мускул-замыкатель, или их два, но один из них значительно меньше другого. В группу Eulamellibranchiata были включены представители с пластинчатыми жабрами. Eulamellibranchiata подразделялись на 4 подотряда: Schizodonta, Heterodonta, Adapedonta и Anomalodesmata[118][119].
Классификация на основе строения замка |
Ниже представлена схема вышеупомянутой классификации двустворчатых, разработанной Ньюэллом в 1965 году и основанной на морфологии замка (знаком † отмечены вымершие таксоны)[110].
Подкласс | Отряд |
|---|---|
Palaeotaxodonta | Nuculoida |
Cryptodonta | † Praecardioida Solemyoida |
Pteriomorphia | Arcoida † Cyrtodontoida Limoida Mytiloida Ostreoida (ранее включались в Pterioida) † Praecardioida Pterioida |
Palaeoheterodonta | Trigonioida (единственный современный род — Neotrigonia) Unionoida † Modiomorpha |
Heterodonta — разнозубые | † Cycloconchidae † Hippuritoida † Lyrodesmatidae Myoida † Redoniidae Veneroida |
Anomalodesmata | Pholadomyoida |
Монофилия подкласса Anomalodesmata является спорной. В настоящее время обычно его относят к подклассу разнозубых[111][114][120].
Классификация на основе строения жабр |
Выше (см. раздел Дыхательная система) приводилась альтернативная классификация двустворчатых моллюсков, основанная на строении жабр[121]. По этому признаку выделяются группы Protobranchia, Filibranchia, Eulamellibranchia и Septibranchia. Первая группа соответствует подклассам Palaeotaxodonta и Cryptodonta Ньюэлла, вторая — Pteriomorphia. Все остальные таксоны по Ньюэллу принадлежат к группе Eulamellibranchia, кроме надсемейства Poromyoidea, соответствующего таксону Septibranchia[122].
Классификация 2010 года |
В мае 2010 года журнал Malacologia опубликовал новую классификацию двустворчатых. При её составлении авторы использовали целый ряд признаков и характеристик: различную филогенетическую информацию, в том числе данные молекулярного анализа, морфологию раковины и её микроструктуру, а также биогеографические, палеобиогеографические и стратиграфические данные. В этой классификации выделяется 324 семейства, из которых 214 известны исключительно по ископаемым останкам, а 110 оставшихся возникли в недавнем прошлом и могли даже не оставить ископаемых следов[123]. Как было упомянуто, в 2012 году она была доработана специалистами WoRMS[124].
Ниже представлена схема этой классификации († обозначены вымершие таксоны)[125]:
Группа Euprotobranchia
Подкласс Разнозубые (Heterodonta) Инфракласс Archiheterodonta
Инфракласс Euheterodonta
| Подкласс Palaeoheterodonta
Подкласс Protobranchia
| Подкласс Pteriomorphia
Инфракласс Eupteriomorphia
|
Разнообразие двустворчатых |
В 2010 году Маркус Губер (англ. Markus Huber) в своей работе Compendium of Bivalves оценил общее число видов двустворчатых в 9200 и объединил их в 106 семейств[3]. Согласно Губеру, часто встречаемая в литературе оценка количества видов Bivalvia в 20 тысяч видов не может быть подтверждена. В нижеследующей таблице представлены основные семейства двустворчатых, а также количество входящих в них родов и видов.
| Подкласс | Надсемейство | Семейства | Роды | Виды |
|---|---|---|---|---|
Heterodonta | 64 (в том числе 1 пресноводное) | &0800800 (в том числе 16 пресноводных) | &56005600 (в том числе 270 пресноводных) | |
Arcticoidea | 2 | &00066 | &001313 | |
Cardioidea | 2 | &003838 | &0260260 | |
Chamoidea | 1 | &00066 | &007070 | |
Clavagelloidea | 1 | &00022 | &002020 | |
Crassatelloidea | 5 | &006565 | &0420420 | |
Cuspidarioidea | 2 | &002020 | &0320320 | |
Cyamioidea | 3 | &002222 | &0140140 | |
Cyrenoidea | 1 | &00066 (3 пресноводных) | &006060 (30 пресноводных) | |
Cyrenoidoidea | 1 | &00011 | &00066 | |
Dreissenoidea | 1 | &00033 (2 пресноводных) | &002020 (12 пресноводных) | |
Galeommatoidea | 4 !около 4 | &0100около 100 | &0500около 500 | |
Gastrochaenoidea | 1 | &00077 | &003030 | |
Glossoidea | 2 | &002020 | &0110110 | |
Hemidonacoidea | 1 | &00011 | &00066 | |
Hiatelloidea | 1 | &00055 | &002525 | |
Limoidea | 1 | &00088 | &0250250 | |
Lucinoidea | 2 | &0085около 85 | &0500около 500 | |
Mactroidea | 4 | &004646 | &0220220 | |
Myoidea | 3 | &001515 (1 пресноводный) | &0130130 (1 пресноводный) | |
Pandoroidea | 7 | &003030 | &0250250 | |
Pholadoidea | 2 | &003434 (1 пресноводный) | &0200200 (3 пресноводных) | |
Pholadomyoidea | 2 | &00033 | &002020 | |
Solenoidea | 2 | &001717 (2 пресноводных) | &0130130 (4 пресноводных) | |
Sphaerioidea | 1 !(1 пресноводный) | &0005(5 пресноводных) | &0200(200 пресноводных) | |
Tellinoidea | 5 | &0110110 (2 пресноводных) | &0900900 (15 пресноводных) | |
Thyasiroidea | 1 | &0012около 12 | &0100около 100 | |
Ungulinoidea | 1 | &001616 | &0100100 | |
Veneroidea | 4 | &0104104 | &0750750 | |
Verticordioidea | 2 | &001616 | &0160160 | |
Palaeoheterodonta | 7 (в том числе 6 пресноводных) | &0171171 (в том числе 170 пресноводных) | &0908908 (в том числе 900 пресноводных) | |
Trigonioidea | 1 | &00011 | &00088 | |
Unionoidea | 6 !(6 пресноводных) | &0170(170 пресноводных) | &0900(900 пресноводных) | |
Protobranchia | 10 | &004949 | &0700700 | |
Manzanelloidea | 1 | &00022 | &002020 | |
Nuculanoidea | 6 | &003232 | &0460460 | |
Nuculoidea | 1 | &00088 | &0170170 | |
Sapretoidea | 1 | &0005около 5 | &001010 | |
Solemyoidea | 1 | &00022 | &003030 | |
Pteriomorpha | 25 | &0240240 (в том числе 2 пресноводных) | &20002000 (в том числе 11 пресноводных) | |
Anomioidea | 2 | &00099 | &003030 | |
Arcoidea | 7 | &006060 (1 пресноводный) | &0570570 (6 пресноводных) | |
Dimyoidea | 1 | &00033 | &001515 | |
Limoidea | 1 | &00088 | &0250250 | |
Mytiloidea | 1 | &005050 (1 пресноводный) | &0400400 (5 пресноводных) | |
Ostreoidea | 2 | &002323 | &008080 | |
Pectinoidea | 4 | &006868 | &0500500 | |
Pinnoidea | 1 | &00033 (+) | &005050 | |
Plicatuloidea | 1 | &00011 | &002020 | |
Pterioidea | 5 | &00099 | &008080 |
Охрана двустворчатых |

Alasmidonta raveneliana — один из видов двустворчатых моллюсков, находящихся на грани вымирания[126]
В Международной Красной книге МСОП приведены по состоянию на октябрь 2013 года данные для 698 видов двустворчатых, из которых 29 видов считаются вымершими, а ещё 164 вида находятся в категориях повышенного риска (категории CR, EN, VU)[127].
В Красную книгу России занесено 34 вида моллюсков, большая часть из которых относится к отряду Unionoida (из них 8 видов рода жемчужница и 7 видов рода миддендорффиная)[128].
Подавляющее большинство видов, находящихся под угрозой исчезновения различной степени риска, являются пресноводными[129]. Для двустворчатых, находящихся под угрозой исчезновения, характерен ряд общих черт: позднее половое созревание, относительно высокая продолжительность жизни, низкая плодовитость, ограниченный ареал, специфическая среда обитания, наличие специфических хозяев для глохидиев (это характерно для унионид)[129].
Взаимодействие с человеком |
Использование человеком |
Человек использует некоторые виды двустворчатых в пищу, как источник материала для украшений (перламутр и жемчуг), в качестве сувениров или даже денег. Нередко двустворчатых применяют для очистки воды. Выделяемый некоторыми двустворчатыми моллюсками биссус используют для изготовления особой ткани — виссона.
- В пищевой промышленности

Устрица съедобная (Ostrea edulis)
Двустворчатые моллюски, например мидии и устрицы, издревле служили пищей человеку[130]. В 2010 году в аквакультурных хозяйствах было выращено 14,2 млн тонн моллюсков, что составляет 23,6 % от всей массы моллюсков, употребляемых в пищу[131]. В 1950 году, когда Продовольственная и сельскохозяйственная организация ООН начала публиковать подобную статистику, общая масса потреблённых двустворчатых моллюсков была оценена в 1 007 419 тонн[132]. В 2000 году эта величина составляла уже 10 293 607, а в 2010 — 14 616 172. В частности, потребление мидий в 2010 году составило 1 901 314 (десятилетием раньше — 1 568 417) тонн, устриц — 4 592 529 (3 858 911) тонн, гребешков — 2 567 981 (1 713 453) тонн[132]. В Китае потребление двустворчатых моллюсков от 1970 к 1997 году выросло в 400 раз[133].
Некоторые страны регулируют импорт двустворчатых и других морепродуктов, в основном для минимизации риска отравления токсинами, которые накапливаются в этих организмах[134].
В настоящее время добыча двустворчатых моллюсков уступает их искусственному разведению в марикультуре. Таким образом на специальных хозяйствах выращивают мидий и устриц. Особенно больших успехов такие хозяйства достигли в США, Японии, Франции, Испании, Италии. В России подобные хозяйства расположены на берегах Чёрного, Белого, Баренцева и Японского морей. Кроме того, в Японии развита марикультура морской жемчужницы (Pinctada)[135].
- Для биологической очистки вод

Saccostrea cucullata
Уже говорилось, что двустворчатые моллюски, живущие в загрязнённых водах, накапливают в своих тканях тяжёлые металлы и стойкие органические загрязнители. Дело в том, что при попадании в моллюска эти вещества не разрушаются ферментами, а накапливаются в его тканях. Эти вещества могут быть вредными как для самих моллюсков, так и людей, поедающих их. Однако это даёт и возможность использовать двустворчатых моллюсков как индикаторов наличия и количества загрязнителей в окружающей среде[136].
Однако для такого использования Bivalvia существуют и ограничения. Уровень загрязнителей в тканях зависит от вида, возраста, размера моллюска, а также времени года и других факторов. Изучение нескольких видов двустворчатых моллюсков, обитающих в лагунах Ганы, дало совершенно аномальные результаты. Для этого есть ряд причин. Например, уровень цинка и железа повышается во время сезона дождей, так как потоки воды смывают их с крыш домов из оцинкованного железа. Уровень кадмия у молодых особей был ниже, чем у старых: хотя в ткани поступал кадмий извне, из-за быстрого роста моллюска уровень кадмия оставался низким[137]. В ходе исследований, проведённых вблизи Владивостока, было показано, что у двустворчатых, живущих в гаванях, содержание тяжёлых металлов понижено, несмотря на повышенную загрязнённость ими воды. Возможно, это объясняется тем, что вода загрязнена не только металлами, но и органикой. Такая вода более питательна, и, как следствие, моллюски снижают фильтрующую активность[138].
Разбитые раковины двустворчатых моллюсков, образующиеся как побочный продукт от консервной промышленности, могут быть использованы для захвата загрязнителей из воды. Было показано, что, если pH воды остаётся щелочным, осколки раковин вбирают кадмий, свинец и другие тяжёлые металлы, замещая на них кальций, входящий в состав арагонита, и переводя их таким образом в твёрдое состояние[139]. Так, устрица Saccostrea cucullata снизила уровень меди и кадмия в водах Персидского залива, причём это достигалось не только за счёт живых моллюсков, но и пустых раковин[140].
- В производстве предметов роскоши и украшений

Ожерелье из жемчуга

Жемчужная ферма в Сераме, Индонезия
Большинство моллюсков, имеющих раковины, образуют жемчуг, но коммерческой ценностью обладают лишь жемчужины, покрытые слоем перламутра. Их создают только двустворчатые и некоторые брюхоногие моллюски[141][142]. Среди натурального жемчуга наибольшей ценностью обладает жемчуг двустворчатых моллюсков Pinctada margaritifera и Pinctada mertensi, обитающих в тропической и субтропической части Тихого океана. Промышленная добыча жемчуга на жемчужных фермах основана на контролируемом внедрении твёрдых частиц в устриц. Материалом для внедряемых частиц часто служат перетёртые раковины других моллюсков. Использование этого материала в промышленных масштабах поставило некоторые пресноводные виды двустворчатых в юго-восточной части США на грань вымирания[142]. Промышленное разведение жемчуга послужило толчком к интенсивному изучению болезней двустворчатых, необходимому для обеспечения здоровья поголовья культивируемых видов[143].

Изготовление пуговиц из раковины двустворчатого моллюска
Добываемый из раковин перламутр используется для изготовления различных изделий, например пуговиц, а также для инкрустаций[144].
Виссон — дорогостоящая ткань, материалом для изготовления которой служит биссус. Это белковый материал, который выделяют двустворчатые моллюски некоторых видов (наиболее известен Pinna nobilis) для крепления к морскому дну[145]. Прокопий Кесарийский, описывая персидские войны середины VI века н. э., утверждал, что только представителям правящих классов дозволялось носить хламиды из виссона[146].
- Прочее использование

Статуэтки из раковин двустворчатых
Раковины двустворчатых нередко служат предметом коллекционирования. Есть много частных и общественных коллекций раковин, крупнейшая из которых находится в Смитсоновском институте и включает около 20 млн образцов[147].
Нередко раковины двустворчатых используются для декорирования. Их спрессовывают в пластины, служащие для украшения стен или садовых дорожек, ими украшают рамки для фотографий, зеркала и другие предметы. Из них часто делают орнаменты. Нередко раковины используют для изготовления украшений. Древнейшие образцы выделанных раковин двустворчатых были обнаружены в пещере в Индонезии, их возраст оценивается в 32 тысячи лет[148].
Индейцы, живущие вблизи восточного побережья, использовали кусочки раковин (например, двустворчатого моллюска Mercenaria mercenaria, а также некоторых брюхоногих) как деньги[149]. Люди племени виннебаго из Висконсина нашли пресноводным двустворчатым многочисленные применения. Так, их раковины служат им в качестве ложек, чаш, ковшей, прочей посуды и инструментов. Они делают из раковин крючки и приманки для ловли рыбы[150].
Измельчённые раковины добавляются в корм для домашних птиц как кальцийсодержащая добавка[151].
Вредители |

Скопления (друзы) дрейссены речной (Dreissena polymorpha)
Корабельные черви поселяются в погружённой в воду древесине, в том числе в подводных частях деревянных лодок и кораблей, а также в стационарных гидротехнических сооружениях. В процессе своей жизнедеятельности (см. раздел Питание) корабельный червь протачивает в древесине многочисленные ходы, чем способствует её быстрому разрушению[152]. Наносимый корабельными червями ежегодный ущерб исчисляется миллионами[153].
Мелкий двустворчатый моллюск Dreissena polymorpha прикрепляется к твёрдому субстрату биссусом и образует значительные скопления. Часто она поселяется в трубах и водоводах, засоряя их[154].
Двустворчатые и здоровье человека |
Давно известно, что потребление сырых или плохо приготовленных двустворчатых моллюсков может приводить к инфекционным заболеваниям. Они могут вызываться как бактериями, изначально обитающими в море (например, возбудитель холеры Vibrio spp.), или же бактериями и вирусами, попадающими в прибрежную зону из сточных вод. Будучи фильтраторами, двустворчатые моллюски пропускают сквозь свои жабры колоссальное количество воды и отфильтровывают не только пищевые частицы, но и микробов. Микробы сохраняются в тканях моллюска и накапливаются в его печени[133][155].
Существуют двустворчатые моллюски, ядовитые для человека; отравление может сопровождаться паралитическим эффектом (англ. Paralytic shellfish poisoning (PSP)), потерей памяти (англ. Amnesic shellfish poisoning (ASP)), гастроэнтеритами, долговременными нейрологическими расстройствами и даже смертью. Ядовитость двустворчатых моллюсков обусловлена накоплением ими выделяющих токсины одноклеточных: диатомовых водорослей или динофлагеллят, которых они отфильтровывают из воды; иногда токсины сохраняются даже в хорошо приготовленных моллюсках[156]. Так, ядовитость двустворчатого моллюска Crassostrea echinata обусловлена токсинами протиста Pyrodinium bahamense из группы динофлагеллят[157].
Гигантская тридакна (Tridacna gigas) теоретически может представлять опасность для человека, во-первых, из-за острых краёв, а во-вторых, она может зажать створками конечность ныряльщика. Однако в настоящий момент случаев человеческих смертей из-за тридакны зарегистрировано не было[158].
Двустворчатые моллюски в культуре |

«Рождение Венеры». Фреска из Помпей

«Рождение Венеры» Боттичелли
Двустворчатые моллюски в древнегреческой культуре были связаны с культом Афродиты. Так, на картине «Рождение Венеры» кисти Боттичелли Венера плывёт к берегу на створке раковины. Раковина присутствует и на фреске с аналогичным сюжетом, найденной в Помпеях. Римляне чтили Венеру и ставили в её честь святилища в своих садах, моля её о дожде и богатом урожае[159], поэтому раковина гребешка и других двустворчатых стала символом плодородия[160]. Раковины морского гребешка служили существенной частью культа богини-матери в Фесте[161]. Изображение морского гребешка, а порой и саму раковину, во времена Средневековья прикрепляли к одежде путешественники, пустившиеся в паломничество к Святым Местам. Подобная раковина служила одновременно и духовным, и совершенно земным целям, как то: сосудом для сбора милостыни, тарелкой для еды. Из-за привычки паломников украшать себя подобным образом появилось современное французское название морского гребешка — «раковина Св. Иакова» (coquille St. Jacques)[162].
Изображение раковины двустворчатого моллюска используется во многих произведениях архитектуры, мебели. Наконец, она является логотипом Royal Dutch Shell, крупной нефтяной и газовой компании[163].
Примечания |
↑ Книпович Н. М. Пластинчатожаберные // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
↑ 1234 Шарова, 2002, с. 304.
↑ 123 Huber, Markus. Compendium of Bivalves. A Full-color Guide to 3,300 of the World's Marine Bivalves. A Status on Bivalvia after 250 Years of Research. — ConchBooks, 2010. — P. 23. — ISBN 978-3-939767-28-2.
↑ Marine Proteins and Peptides: Biological Activities and Applications / Se-Kwon Kim (Ed.). — John Wiley & Sons, 2013. — P. 41. — ISBN 9781118375105.
↑ Данные о числе видов для России и всего мира на сайте Зоологического института РАН (неопр.). Проверено 12 января 2014.
↑ Linnaeus, Carolus. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata : . — Laurentii Salvii, 1758. — P. 645.
↑ 12 Bivalve (неопр.). McGraw-Hill Dictionary of Scientific and Technical Terms. McGraw-Hill Companies. Проверено 7 мая 2012.
↑ Le Renard, J.; Sabelli, B.; Taviani, M. (1996). “On Candinia (Sacoglossa: Juliidae), a new fossil genus of bivalved gastropods”. Journal of Paleontology. 70 (2): 230—235. JSTOR 1306386..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
↑ The Phylum Brachiopoda (неопр.). Earthlife Web. Проверено 5 мая 2012.
↑ Ostracoda (неопр.). Oxford Dictionaries. Oxford University Press. Проверено 1 июля 2012.
↑ Webb, J. (1979). “A reappraisal of the palaeoecology of conchostracans (Crustacea: Branchiopoda)”. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 158 (2): 259—275.
↑ Севергин В. М. Опыт минералогическаго землеописания Российского государства. Ч. 1. СПб.: 1809. XYIII, 262 c.
↑ Yonge, C. M. The Sea Shore. — Collins, 1949. — P. 228.
↑ Bivalves (неопр.). Arctic Ocean Diversity. Проверено 21 апреля 2012.
↑ Adamussium colbecki (Smith, 1902) (неопр.) (недоступная ссылка). Antarctic Field Guide. Проверено 21 апреля 2012. Архивировано 14 октября 2013 года.
↑ 123 Extreme Bivalves (англ.) (недоступная ссылка). Rock the Future. Paleontological Research Institution and its Museum of the Earth. Проверено 14 октября 2013. Архивировано 8 января 2014 года.
↑ Rice, Tony. Hydrothermal vents (неопр.). Deep Ocean. Fathom. Проверено 21 апреля 2012. Архивировано 10 декабря 2008 года.
↑ Christian, A. D. Life History and Population Biology of the State Special Concern Ouachita Creekshell, Villosa arkansasensis (I. Lea 1862) (неопр.). Arkansas State University (2007). Проверено 21 апреля 2012.
↑ Karatayev, A. Y.; Burlakova, L. E.; Karatayev, V. A.; Boltovskoy, D. (2010). “Limnoperna fortunei versus Dreissena polymorpha: population densities and benthic community impacts of two invasive freshwater bivalves”. Journal of Shellfish Research. 29 (4): 975—984. DOI:10.2983/035.029.0432.
↑ Hoddle, M. S. Quagga and Zebra Mussels (неопр.). Center for Invasive Species Research, UC Riverside (13 Oct 2011). Проверено 21 апреля 2012.
↑ 12345678910 Шарова, 2002, с. 306.
↑ Book review: Conchologists of America (неопр.) (недоступная ссылка). Проверено 19 апреля 2012. Архивировано 7 августа 2012 года.
↑ Kauffman, E. G.; Harries, P. J.; Meyer, C.; Villamil, T.; Arango, C.; Jaecks, G. (2007). “Paleoecology of giant Inoceramidae (Platyceramus) on a Santonian (Cretaceous) seafloor in Colorado”. Journal of Paleontology. 81 (1): 64—81. DOI:10.1666/0022-3360(2007)81[64:POGIPO]2.0.CO;2.
↑ Edmondson, C. H. (1962). “Teredinidae, ocean travelers” (PDF). Occasional Papers of Bernice P. Bishop Museum. 23 (3): 45—59.
↑ 123 Догель, 1981, с. 473.
↑ 123456 Тип Моллюски (неопр.) (недоступная ссылка). Архивировано 7 января 2014 года.
↑ 123 Wells, Roger M. Class Bivalvia (неопр.) (недоступная ссылка). Invertebrate Paleontology Tutorial. State University of New York College at Cortland (1998). Проверено 11 апреля 2012. Архивировано 28 февраля 2010 года.
↑ 12 Kennedy, W. J.; Taylor, J. D.; Hall, A. (1969). “Environmental and biological controls on bivalve shell mineralogy”. Biological Reviews. 4 (4): 499—530. DOI:10.1111/j.1469-185X.1969.tb00610.x.
↑ 123 Bivalvia (неопр.). SpringerImages. Проверено 6 мая 2012.
↑ 12 Barrett, John; Yonge, C. M. Collins Pocket Guide to the Sea Shore. — London : William Collins Sons and Co. Ltd, 1958. — P. 148.
↑ 123 Шарова, 2002, с. 308.
↑ Morton, Brian. Bivalve: The mantle and musculature (неопр.). Encyclopædia Britannica. Проверено 5 мая 2012.
↑ 1234 Двустворчатые моллюски — статья из Биологического энциклопедического словаря
↑ Догель, 1981, с. 476—477.
↑ Dorit, Robert L. Zoology. — Saunders College Publishing, 1991. — P. 679. — ISBN 978-0-03-030504-7.
↑ Шарова, 2002, с. 308—309.
↑ 123 Догель, 1981, с. 477.
↑ Morton, Brian. Bivalve: The digestive system and nutrition (неопр.). Encyclopædia Britannica. Проверено 7 мая 2012.
↑ 123 Bourquin, Avril. Class Bivalvia (Pelecypoda) (неопр.) (недоступная ссылка). The Phylum Mollusca (2000). Проверено 5 мая 2012. Архивировано 5 мая 2012 года.
↑ Шарова, 2002, с. 309.
↑ 12 Cofrancesco, Alfred F. Nervous System and Sense Organs in Bivalves (неопр.). Zebra Mussel Research Program (2002). Проверено 5 мая 2012. Архивировано 15 апреля 2012 года.
↑ Morton, Brian. Mollusk: The nervous system and organs of sensation (неопр.). Encyclopædia Britannica. Проверено 8 июля 2012.
↑ 12 Шарова, 2002, с. 310.
↑ Догель, 1981, с. 477—478.
↑ Allen, J. A.; Morgan, Rhona E. (1981). “The functional morphology of Atlantic deep water species of the families Cuspidariidae and Poromyidae (Bivalvia): an analysis of the evolution of the septibranch condition”. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 294 (1073): 413—546. DOI:10.1098/rstb.1981.0117.
↑ Morton, B. The evolution of eyes in the Bivalvia: new insights // American Malacological Bulletin. — 2008. — Vol. 26, № 1—2. — P. 35—45. — DOI:10.4003/006.026.0205.
↑ Colicchia, G.; Waltner, C.; Hopf, M.; Wiesner, H. (2009). “The scallop's eye — a concave mirror in the context of biology”. Physics Education. 44 (2): 175—179. DOI:10.1088/0031-9120/44/2/009.
↑ 12 Догель, 1981, с. 478.
↑ Шарова, 2002, с. 310—311.
↑ 12 Morton, Brian. Bivalve: The respiratory system (неопр.). Encyclopædia Britannica. Проверено 8 июля 2012.
↑ 12345 Шарова, 2002, с. 311.
↑ 1234 Догель, 1981, с. 479.
↑ Dorit, Robert L. Zoology. — Saunders College Publishing, 1991. — P. 674. — ISBN 978-0-03-030504-7.
↑ 12 Vaughan, Burton. The Bivalve, Poromya granulata (неопр.). Archerd Shell Collection (2008). Проверено 3 апреля 2012.
↑ Morton, Brian. Bivalve: The excretory system (неопр.). Encyclopædia Britannica. Проверено 7 мая 2012.
↑ Пойкилосмотические животные — статья из Большой советской энциклопедии.
↑ 12 Рупперт, Фокс, Барнс, 2008, с. 249.
↑ G. Bello, P. Paparella, A. Corriero, N. Santamaria. Protandric hermaphroditism in the bivalve Arca noae (Mollusca: Arcidae) // Mediterranean Marine Science. — 2013. — Vol. 14, № 1. — P. 86—91.
↑ 1234 Догель, 1981, с. 480.
↑ 1234 Шарова, 2002, с. 312.
↑ Foighil, D. O. (1989). “Planktotrophic larval development is associated with a restricted geographic range in Lasaea, a genus of brooding, hermaphroditic bivalves”. Marine Biology. 103 (3): 349—358. DOI:10.1007/BF00397269.
↑ Helm, M. M.; Bourne, N.; Lovatelli, A. Gonadal development and spawning (неопр.). Hatchery culture of bivalves: a practical manual. FAO (2004). Проверено 8 мая 2012.
↑ Dorit, Robert L. Zoology. — Saunders College Publishing, 1991. — P. 774. — ISBN 978-0-03-030504-7.
↑ 12 Barnes, R. S. K. The Invertebrates: A New Synthesis. — Blackwell Scientific Publications, 1988. — P. 140. — ISBN 978-0-632-03125-2.
↑ 12 Piper, Ross. Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. — Greenwood Press, 2007. — P. 224—225. — ISBN 978-0-313-33922-6.
↑ Honkoop, P. J. C.; Van der Meer, J.; Beukema, J. J.; Kwast, D. (1999). “Reproductive investment in the intertidal bivalve Macoma balthica” (PDF). Journal of Sea Research. 41 (3): 203—212. DOI:10.1016/S1385-1101(98)00053-7.
↑ 12 Butler, P. G. et al. Variability of marine climate on the North Icelandic Shelf in a 1357-year proxy archive based on growth increments in the bivalve Arctica islandica (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2013. — Vol. 373. — P. 141—151. — DOI:10.1016/j.palaeo.2012.01.016.
↑ 12 I. D. Ridgway, C. A. Richardson, S. N. Austad. Maximum Shell Size, Growth Rate, and Maturation Age Correlate With Longevity in Bivalve Molluscs // J Gerontol A Biol Sci Med Sci. — 2011. — Vol. 66А, № 2. — DOI:10.1093/gerona/glq172.
↑ I. Thompson, D. S. Jones, J. W. Ropes. Advanced age for sexual maturity in the ocean quahog Arctica islandica (Mollusca: Bivalvia) // Marine Biology. — 1980. — Vol. 57, № 1. — P. 35—39. — ISSN 1432-1793. — DOI:10.1007/BF00420965.
↑ 12 Шарова, 2002, с. 316.
↑ Lützen, J.; Berland, B.; Bristow, G. A. (2011). “Morphology of an endosymbiotic bivalve, Entovalva nhatrangensis (Bristow, Berland, Schander & Vo, 2010) (Galeommatoidea)” (PDF). Molluscan Research. 31 (2): 114—124.
↑ Солемии — статья из Биологического энциклопедического словаря
↑ Пименова, Пименов, 2005, с. 135.
↑ 12 Barnes, R. S. K. The Invertebrates: A New Synthesis. — Blackwell Scientific Publications, 1988. — P. 132—134. — ISBN 978-0-632-03125-2.
↑ Barnes, R. S. K. The Invertebrates: A New Synthesis. — Blackwell Scientific Publications, 1988. — P. 265. — ISBN 978-0-632-03125-2.
↑ 123 Догель, 1981, с. 481.
↑ Carefoot, Tom. Learn about whelks and relatives: foods, feeding and growth (неопр.) (недоступная ссылка). A snail's odyssey (2010). Проверено 19 апреля 2012. Архивировано 5 июля 2012 года.
↑ 12 Harper, Elizabeth M. (1990). “The role of predation in the evolution of cementation in bivalves” (PDF). Palaeontology. 34 (2): 455—460. (недоступная ссылка)
↑ Bishop, M. J.; Garis, H. (1976). “A note on population densities of mollusca in the River Great Ouse at Ely, Cambridgeshire”. Hydrobiologia. 48 (3): 195—197. DOI:10.1007/BF00028690.
↑ 12 Шарова, 2002, с. 315.
↑ Пименова, Пименов, 2005, с. 134.
↑ Mollusks: Bioluminescence (неопр.).
↑ 12 Шарова, 2002, с. 318.
↑ Шарова, 2002, с. 318—319.
↑ Limaria fragilis (неопр.). Saltcorner. Проверено 20 апреля 2012.
↑ Thorp, J. H.; Delong, M. D.; Casper1, A. F. (1998). “In situ experiments on predatory regulation of a bivalve mollusc (Dreissena polymorpha) in the Mississippi and Ohio Rivers”. Freshwater Biology. 39 (4): 649—661. DOI:10.1046/j.1365-2427.1998.00313.x.
↑ Hulscher, J. B. (1982). “The oystercatcher Haematopus ostralegus as a predator of the bivalve Macoma balthica in the Dutch Wadden Sea” (PDF). Ardea. 70: 89—152. (недоступная ссылка)
↑ Ingolfsson, Agnar; Bruce T. Estrella (1978). “The development of shell-cracking behavior in herring gulls” (PDF). The Auk. 95 (3): 577—579. Используется устаревший параметр|coauthors=(справка)
↑ Hall, K. R. L.; Schaller, G. B. (1964). “Tool-using behavior of the California sea otter”. Journal of Mammalogy. 45 (2): 287—298. DOI:10.2307/1376994. JSTOR 1376994.
↑ Fukuyamaa, A. K.; Olivera, J. S. (1985). “Sea star and walrus predation on bivalves in Norton Sound, Bering Sea, Alaska”. Ophelia. 24 (1): 17—36. DOI:10.1080/00785236.1985.10426616.
↑ Wodinsky, Jerome (1969). “Penetration of the shell and feeding on gastropods by octopus”. American Zoologist. 9 (3): 997—1010. DOI:10.1093/icb/9.3.997.
↑ Моллюски — статья из Энциклопедии Кольера
↑ Pacific razor clam (неопр.). California Department of Fish and Game (2001). Проверено 9 мая 2012. Архивировано 24 августа 2013 года.
↑ Bourquin, Avril. Bivalvia: The foot and locomotion (неопр.). The Phylum Mollusca (2000). Проверено 19 апреля 2012. Архивировано 24 августа 2013 года.
↑ Hodgson, A. N. (1981). “Studies on wound healing, and an estimation of the rate of regeneration, of the siphon of Scrobicularia plana (da Costa)”. Journal of Experimental Marine Biology and Ecology. 62 (2): 117—128. DOI:10.1016/0022-0981(82)90086-7.
↑ Fleming, P. A.; Muller, D.; Bateman, P. W. (2007). “Leave it all behind: a taxonomic perspective of autotomy in invertebrates”. Biological Reviews. 82 (3): 481—510. DOI:10.1111/j.1469-185X.2007.00020.x. PMID 17624964.
↑ Metzger Michael J., Villalba Antonio, Carballal María J., Iglesias David, Sherry James, Reinisch Carol, Muttray Annette F., Baldwin Susan A., Goff Stephen P. Widespread transmission of independent cancer lineages within multiple bivalve species // Nature. — 2016. — Июнь (т. 534, № 7609). — С. 705—709. — ISSN 0028-0836. — DOI:10.1038/nature18599.
↑ Mark A. Farmer, William K. Fitt, Robert K. Trench. Morphology of the Symbiosis Between Corculum cardissa (Mollusca: Bivalvia) and Symbiodinium corculorum (Dinophyceae) // Biol. Bull. — 2001. — Vol. 200. — P. 336—343.
↑ Пименова, Пименов, 2005, с. 131.
↑ Campbell, N. A. Biology, Sixth Edition. — Benjamin Cummings, 2001. — P. 643. — ISBN 978-0-201-75054-6.
↑ Pojeta, J. (2000). “Cambrian Pelecypoda (Mollusca)”. American Malacological Bulletin. 15: 157—166.
↑ Schneider, J. A. (November 2001). “Bivalve systematics during the 20th century”. Journal of Paleontology. 75 (6): 1119—1127. DOI:10.1666/0022-3360(2001)075<1119:BSDTC>2.0.CO;2. Проверено 2008-10-05.
↑ Gubanov, A. P., Kouchinsky, A. V. and Peel, J. S. (2007). “The first evolutionary-adaptive lineage within fossil molluscs”. Lethaia. 32 (2): 155—157. DOI:10.1111/j.1502-3931.1999.tb00534.x.
↑ Gubanov, A. P., and Peel, J. S. (2003). “The early Cambrian helcionelloid mollusc Anabarella Vostokova”. Palaeontology. 46 (5): 1073—1087. DOI:10.1111/1475-4983.00334.
↑ Zong-Jie, F. (2006). “An introduction to Ordovician bivalves of southern China, with a discussion of the early evolution of the Bivalvia”. Geological Journal. 41 (3—4): 303—328. DOI:10.1002/gj.1048.
↑ 12 Fossil Record (неопр.). University of Bristol. Проверено 11 мая 2012.
↑ Brosius, L. Fossil Brachiopods (неопр.) (недоступная ссылка). GeoKansas. Kansas Geological Survey (2008). Проверено 2 июля 2012. Архивировано 5 июля 2008 года.
↑ Gould, Stephen; C. Bradford Calloway (Autumn, 1980). “Clams and brachiopods — ships that pass in the night”. Paleobiology. 6 (4): 383—396. JSTOR 2400538. Используется устаревший параметр|coauthors=(справка); Проверьте дату в|date=(справка на английском)
↑ Moore, R. C. Treatise on Invertebrate Paleontology, Part N: Mollusca 6, Bivalvia (Vol. 3). — Geological Society of America, 1969. — P. 275. — ISBN 978-0-8137-3026-4.
↑ 12 Newell, Norman D. Bivalvia Systematics // Treatise on Invertebrate Paleontology Part N. — The Palentological Institute, 1969. — ISBN 978-0-8137-3014-1.
↑ 12 Giribet G., Wheeler W. (2002). “On bivalve phylogeny: a high-level analysis of the Bivalvia (Mollusca) based on combined morphology and DNA sequence data”. Invertebrate Biology. 121 (4): 271—324. DOI:10.1111/j.1744-7410.2002.tb00132.x.
↑ Bieler R., Mikkelsen P. M. (2006). “Bivalvia — a look at the branches”. Zoological Journal of the Linnean Society. 148 (3): 223—235. DOI:10.1111/j.1096-3642.2006.00255.x.
↑ Mikkelsen, P. M.; Bieler, R.; Kappner, I.; Rawlings, T. A. (2006). “Phylogeny of Veneroidea (Mollusca: Bivalvia) based on morphology and molecules”. Zoological Journal of the Linnean Society. 148 (3): 439—521. DOI:10.1111/j.1096-3642.2006.00262.x.
↑ 12 Taylor, John D.; Williams, Suzanne T.; Glover, Emily A.; Dyal, Patricia. A molecular phylogeny of heterodont bivalves (Mollusca: Bivalvia: Heterodonta): New analyses of 18S and 28S rRNA genes // Zoologica Scripta. — 2007. — Vol. 36, № 6. — P. 587—606. — DOI:10.1111/j.1463-6409.2007.00299.x.
↑ Taylor, John D.; Glover, Emily A.; Williams, Suzanne T. (2009). “Phylogenetic position of the bivalve family Cyrenoididae — removal from (and further dismantling of) the superfamily Lucinoidea”. Nautilus. 123 (1): 9—13.
↑ Tëmkin, I. (2010). “Molecular phylogeny of pearl oysters and their relatives (Mollusca, Bivalvia, Pterioidea)” (PDF). BMC Evolutionary Biology. 10: 342. DOI:10.1186/1471-2148-10-342.
↑ Taylor, John D.; Glover, Emily A.; Smith, Lisa; Dyal, Patricia; Williams, Suzanne T. (September 2011). “Molecular phylogeny and classification of the chemosymbiotic bivalve family Lucinidae (Mollusca: Bivalvia)”. Zoological Journal of the Linnean Society. 163 (1): 15—49. DOI:10.1111/j.1096-3642.2011.00700.x. Используется устаревший параметр|month=(справка); Неизвестный параметр|doi_brokendate=(справка)
↑ Schneider, Jay A. (2001). “Bivalve systematics during the 20th century”. Journal of Paleontology. 75 (6): 1119—1127. DOI:10.1666/0022-3360(2001)075<1119:BSDTC>2.0.CO;2. ISSN 0022-3360.
↑ Ponder, W. F.; Lindberg, David R. Phylogeny and Evolution of the Mollusca. — University of California Press, 2008. — P. 117. — ISBN 978-0-520-25092-5.
↑ Harper, E. M.; Dreyer, H.; Steiner, G. (2006). “Reconstructing the Anomalodesmata (Mollusca: Bivalvia): morphology and molecules”. Zoological Journal of the Linnean Society. 148 (3): 395—420. DOI:10.1111/j.1096-3642.2006.00260.x.
↑ Franc, A. Classe de Bivalves // Traité de Zoologie: Anatomie, Systématique, Biologie : . — Masson et Cie, 1960. — Vol. 5. — P. 1845—2164.
↑ Septibranchia (неопр.). McGraw-Hill Dictionary of Scientific and Technical Terms. McGraw-Hill Companies. Проверено 7 мая 2012.
↑ Bouchet, Philippe; Rocroi, Jean-Pierre; Bieler, Rüdiger; Carter, Joseph G.; Coan, Eugene V. Nomenclator of Bivalve Families with a Classification of Bivalve Families // Malacologia. — 2010. — Vol. 52, № 2. — P. 1—184. — DOI:10.4002/040.052.0201.
↑ WoRMS: Bivalvia (неопр.).
↑ Carter, J. G.; Altaba, C. R.; Anderson, L. C.; Araujo, R.; Biakov, A. S.; Bogan, A. E.; Campbell, D. C.; Campbell, M.; Chen, J.; Cope, J. C. W.; Delvene. G.; Dijkstra, H. H.; Fang, Z.; Gardner, R. N.; Gavrilova, V. A.; Goncharova, I. A.; Harries, P. J.; Hartman, J. H.; Hautmann, M.; Hoeh, W. R.; Hylleberg, J.; Jiang, B.; Johnston, P.; Kirkendale, L.; Kleemann, K.; Koppka, J.; Kříž, J.; Machado, D.; Malchus, N.; Márquez-Aliaga, A.; Masse, J-P.; McRoberts, C. A.; Middelfart, P. U.; Mitchell, S.; Nevesskaja, L. A.; Özer, S.; Pojeta, J. Jr.; Polubotko, I. V.; Pons, J. M.; Popov, S.; Sánchez, T.; Sartori, A. F.; Scott, R. W.; Sey, I. I.; Signorelli, J. H.; Silantiev, V. V.; Skelton, P. W.; Steuber, T.; Waterhouse, J. B.; Wingard, G. L.; Yancey, T. (2011). “A synoptical classification of the Bivalvia (Mollusca)” (PDF). Paleontological Contributions. 4: 1—47. Используется устаревший параметр|coauthors=(справка)
↑ Alasmidonta raveneliana (англ.). The IUCN Red List of Threatened Species.
↑ IUCN Red List of Threatened Species. Version 2013.1 (неопр.). IUCN. Проверено 24 октября 2013. Архивировано 16 августа 2013 года.
↑ Красная книга России: моллюски (неопр.). Проверено 19 августа 2013.
↑ 12 The Conservation Biology of Molluscs // E. Alison Kay. — International Union for Conservation of Nature and Natural Resources, 1995. — ISBN 2-8317-0053-1.
↑ Mannino, M. A., and Thomas, K. D. (February 2002). “Depletion of a resource? The impact of prehistoric human foraging on intertidal mollusc communities and its significance for human settlement, mobility and dispersal”. World Archaeology. 33 (3): 452—474. DOI:10.1080/00438240120107477.
↑ FAO 2010: Fishery and Aquaculture Statistics (неопр.).
↑ 12 Fishery Statistical Collections: Global Production (неопр.). FAO Fisheries and Aquaculture Department. Проверено 23 мая 2012.
↑ 12 Potasman, I.; Paz, A.; Odeh, M. (2002). “Infectious outbreaks associated with bivalve shellfish consumption: a worldwide perspective”. Clinical Infectious Diseases. 35 (8): 921—928. DOI:10.1086/342330. PMID 12355378.
↑ Importing fishery products or bivalve molluscs (неопр.). United Kingdom: Food Standards Agency. Проверено 2 октября 2008. Архивировано 19 ноября 2012 года.
↑ Шарова, 2002, с. 317.
↑ Burns, K. A.; Smith, J. L. (1981). “Biological monitoring of ambient water quality: the case for using bivalves as sentinel organisms for monitoring petroleum pollution in coastal waters”. Estuarine, Coastal and Shelf Science. 30 (4): 433—443. DOI:10.1016/S0302-3524(81)80039-4.
↑ Otchere, F. A. (2003). “Heavy metals concentrations and burden in the bivalves (Anadara (Senilia) senilis, Crassostrea tulipa and Perna perna) from lagoons in Ghana: model to describe mechanism of accumulation/excretion”. African Journal of Biotechnology. 2 (9): 280—287.
↑ Shulkin, V. M.; Kavun, V. I. A. The use of marine bivalves in heavy metal monitoring near Vladivostok, Russia // Marine Pollution Bulletin. — 1995. — Vol. 31, № 4—12. — P. 330—333. — DOI:10.1016/0025-326X(95)00169-N.
↑ Reilly, Michael. Sea Shells Used to Clean Up Heavy Metals (27 апреля 2009). Проверено 18 мая 2012.
↑ Azarbad, H.; Khoi, A. J.; Mirvaghefi, A.; Danekar, A.; Shapoori, M. (2010). “Biosorption and bioaccumulation of heavy metals by rock oyster Saccostrea cucullata in the Persian Gulf” (PDF). International Aquatic Research. 2010 (2): 61—69. ISSN 2008-4935. Архивировано из оригинала (PDF) 2013-10-17. Проверено 2013-10-29.
↑ Ruppert, Fox, Barnes, 2004, pp. 300—343.
↑ 12 Ruppert, Fox, Barnes, 2004, pp. 367—403.
↑ Jones, J. B., and Creeper, J. (April 2006). “Diseases of Pearl Oysters and Other Molluscs: a Western Australian Perspective”. Journal of Shellfish Research. 25 (1): 233—238. DOI:10.2983/0730-8000(2006)25[233:DOPOAO]2.0.CO;2.
↑ Раковина / Иванов А. В. // Проба — Ременсы. — М. : Советская энциклопедия, 1975. — С. 447. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 21).
↑ Webster’s Third New International Dictionary (Unabridged) 1976. G. & C. Merriam Co., p. 307.
↑ Turner, R. D., and Rosewater, J. (June 1958). “The Family Pinnidae in the Western Atlantic”. Johnsonia. 3 (38): 294.
↑ The magnificent shells of the Smithsonian (неопр.). Smithsonian.com. Проверено 5 мая 2012.
↑ Catling, Chris. Shell Tools Rewrite Australasian Prehistory (6 июля 2009). Архивировано 17 марта 2013 года. Проверено 18 мая 2012.
↑ Dubin, Lois Sherr. North American Indian Jewelry and Adornment: From Prehistory to the Present. — Harry N. Abrams, 1999. — P. 170—171. — ISBN 978-0-8109-3689-8.
↑ Kuhm, H. W. (2007). “Aboriginal uses of shell”. The Wisconsin Archeologist. 17 (1): 1—8.
↑ Poultry Grit, Oystershell and Wood Shavings (неопр.). Ascott Smallholding Supplies Ltd. Проверено 18 мая 2012.
↑ Корабельный червь — статья из Большой советской энциклопедии.
↑ Карпов Н. Корабельный червь (рус.). «Земля и люди», январь — март (2009). Проверено 22 августа 2013. Архивировано 24 августа 2013 года.
↑ Шарова, 2002, с. 319.
↑ Rippey, S. R. (1994). “Infectious diseases associated with molluscan shellfish consumption”. Clinical Microbiology. 7 (4): 419—425. DOI:10.1128/CMR.7.4.419.
↑ Wekell, John C.; Hurst, John; Lefebvre, Kathi A. (2004). “The origin of the regulatory limits for PSP and ASP toxins in shellfish” (PDF). Journal of Shellfish Research. 23 (3): 927—930. (недоступная ссылка)
↑ J. L. Maclean. Paralytic Shellfish Poison in Various Bivalves, Port Moresby, 1973 // Pacific Science. — 1975. — Vol. 29, № 4. — P. 349—352.
↑ Sergey M. Govorushko. Natural Processes and Human Impacts: Interactions between Humanity and the Environment. — Springer, 2012. — P. 294. — 678 p. — ISBN 978-94-007-1423-6.
↑ Gilmer, Maureen. Venus honored in Roman garden shrines (19 марта 2004). Архивировано 11 мая 2013 года. Проверено 21 мая 2012.
↑ Fontana, D. The Secret Language of Symbols: A Visual Key to Symbols and Their Meanings. — Chronicle Books, 1994. — P. 88, 103. — ISBN 978-0-8118-0462-2.
↑ Мифология — Морская раковина: греческая мифология
↑ Fulcanelli et la façade du palais Jacques Coeur (фр.). Fulcanelli, La rue de l'alchimie à travers l'architecture, les livres et les alchimistes. Проверено 14 июня 2012.
↑ The Shell global homepage (неопр.). Проверено 21 мая 2012.
Литература |
На русском языке |
- Шарова И. Х. Зоология беспозвоночных. — М.: Владос, 2002. — 592 с. — ISBN 5-691-00332-1.
- Догель В. А. Зоология беспозвоночных. 7-е изд. — М.: Высшая школа, 1981. — 614 с.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Зоология беспозвоночных: функциональные и эволюционные аспекты. — М.: Издательский центр «Академия», 2008. — Т. 2. — 448 с. — ISBN 978-5-7695-3495-9.
- Пименова И. Н., Пименов А. В. Зоология беспозвоночных. Теория. Задания. Ответы. — Саратов: Лицей, 2005. — 288 с. — ISBN 5-8053-0308-6.
- Евсеев Г. А., Яковлев Ю. М. Двустворчатые моллюски дальневосточных морей России. — Владивосток: Институт биологии моря, 2006. — 120 с.
- Атлас двустворчатых моллюсков дальневосточных морей России / С. Е. Поздняков. — Владивосток: Дюма, 2000. — 168 с.
На английском языке |
Schneider, Jay A. (2001). “Bivalve systematics during the 20th century”. Journal of Paleontology. 75 (6): 1119—1127. DOI:10.1666/0022-3360(2001)075<1119:BSDTC>2.0.CO;2.
Poutiers J.-M., Bernard R. D. Carnivorous bivalve molluscs (Anomalodesmata) from the tropical western Pacific Ocean, with a proposed classification and a catalogue of recent species // Résultats des Campagnes Musorstom / Ed. by P. Bouchet. — 7th ed. — Brooks / Cole, 1995. — (Mémoires Muséum National d'Histoire Naturelle, vol. 167). — ISBN 0-03-025982-7. — P. 107—188.- Ruppert E. E., Fox R. S., Barnes R. D. Invertebrate Zoology. 7th ed. — Brooks / Cole, 2004. — ISBN 0-03-025982-7.
- Vaught K. C. A Classification of the Living Mollusca. — American Malacologists, 1989. — ISBN 978-0-915826-22-3.
Эта статья входит в число избранных статей русскоязычного раздела Википедии. |
Словари и энциклопедии | Большая российская · Britannica (онлайн) |
|---|---|
| Нормативный контроль | GND: 4040785-8 |


